
文摘
分工发生在广泛的生物。然而,分工是如何出现在没有预先存在的个人间的差异了解甚少。使用一个简单的但是现实的模型,我们表明,在最初的一组相同的个人,分工出现自发如果返回觅食者与其他小组成员分享他们的资源的一部分。在缺乏资源共享的情况下,个体遵循一个活动安排觅食和其他任务之间的交替。non-foraging个人如果美联储的其他个体,他们的交流活动时间表变得中断,导致任务专业化和劳动分工的出现。此外,营养强化分工也存在个体差异。这种差异可能是由于增加代谢率在觅食或主导地位在资源共享的交互。我们的模型提出了一个合理的机制自我组织分工的出现最初在动物组完全相同的个体。这种机制也可以扮演一个角色分工的出现在主要的进化过渡到真社会性和多细胞生物。
介绍
labour-within-individual一致性,但在个人分工变化任务中选择一组的成员1是一个关键的方面的社会生活在人类和动物的社会。分工发生在广泛的生物2,3。例如,在群居昆虫,工人专攻觅食,保卫巢穴,或护理窝4。在某些种类的鸟类,如嘈杂的矿工,一些帮手专攻小鸡供应或侵扰巢捕食者5。在坦噶尼喀湖丽鱼科鱼公主,助手参与捕食者辩护,负责维修或鸡蛋6,7。在骄傲的狮子,个人执行不同的角色在狩猎和领土防卫,他们可以专门照顾宝宝8。
了解社会生活的核心问题是因此这样的分工如何产生。分工的出现通常是模仿的反应阈值模型1,9,10,11,12,13,14。这些模型假设个体不同阈值,确定他们的可能性开始执行任务时感知刺激的任务。个体与低反应阈值更容易承担的任务,从而降低了任务的刺激;这可以防止阈值较高的其他个人承担的任务。因此,个人专注于这些任务,他们有一个较低的阈值。然而,任务专业化和劳动分工只发生如果有预先存在的个人间的差异响应阈值15。尚知之甚少如何自我组织分工可以出现在同质组高度相似甚至相同的个体。
在这里,我们提出一个模型来分工的出现在动物组织相同的个体。模型是现实的假设个体的营养水平下降随着时间的推移,和低营养水平触发觅食16,17,18,19,20.,21,22(无花果。1)。在缺乏个人间的相互作用,一个期望,个人按顺序执行non-foraging行为(此后称为护理),当营养含量高和觅食行为当营养水平较低。然而,群体内部的动物,个人互动和资源共享经常发生23,24。这可能导致一定程度的专业化觅食和non-foraging行为,觅食,给他们资源的一部分可能很快又有觅食,而共享资源的接收器可以推迟觅食。因此,我们认为是各种资源共享场景(图。1 b),为了研究这种常见的机制是否足以产生高度的专业化和劳动分工的任务。

一个我们的模型假设个人可以在两种状态之间切换(实心箭头):觅食(绿色)和护理(蓝色)。觅食时,个人从环境中获取资源(黑点)和存储这些资源在一个临时存储器官(灰色圆圈)。只要资源储存器官,它们在原则上可以与其他个体共享。一旦融入个人的身体(绿点在觅食的人;蓝点在护理个人)贡献个人的营养水平,可以不再被共享。觅食是引发的低营养水平,觅食个人平均比个人护理营养水平较低。b四个资源共享方案考虑在我们的研究中:(1)没有共享:资源不共享;觅食的人消耗掉所有资源本身。(2)平等共享:觅食个人之间共享收集到的资源同样自己和个人护理。(3)Dominance-based共享:每个人都有一个预先分配的主导地位价值和资源共享与这些主导价值观:占统治地位的个人获得更大比例的收集资源。(4)以营养为基础的共享:在(3),但现在主导地位的个体并不是预先分配和常数,但个人的营养水平成正比。
结果
资源共享是充分的劳动分工的出现
我们跑20为每个资源共享复制模拟场景。我们量化分工的程度,出现在模拟使用公制D由杜阿尔特介绍等。25(见方法对细节和无花果。S1在补充其他分工指标)。本文考虑的场景(见方法),D范围从−1 + 1,−1表明严格的任务之间的交替,0表示随机任务之间切换,+ 1表示任务专业化25。图2表明,在缺乏资源共享(无花果。1 b;(1)没有共享),分工度量D呈现出极端值−1,这意味着个人遵循一个活动安排任务之间的交替。护理个人开始饲料为自己获取资源保留所有的资源。连续,他们直到护士营养水平已经下降到一个阈值,诱发他们重新开始觅食。如无花果所示。2 b,结果是非常不同的觅食和护理个人之间共享资源(无花果。1 b;(2)平等分享)。现在的度规D达到积极的价值高于0.5,表明个人专门觅食或护理的时间长。

一个没有共享。在缺乏资源共享的情况下,D=−1这表明个人轮流护理和觅食。觅食是由营养水平下降,个人护理营养水平高于觅食。b平等的分享。如果觅食的人分享他们的收集资源同样与护理的人,D≈0.6代表一个中级水平分工。每个灰色点(左面板)是分工指标从一个复制模拟(n= 20)。蓝色(护理)和绿色(觅食)点(右面板)代表营养水平的所有个体的复制模拟。
分工是钢筋如果觅食者有更高的代谢率
随着代谢率可以不同的任务26,27,我们测试了这些差异如何影响分工的出现。即使与护理相关的代谢率(90%或95%)仅略低于相关的代谢率觅食,分工强烈钢筋(无花果。3),导致营养水平(图的双峰分布。3 b)。如果与护理相关代谢率高于觅食,分工仍会发生的,但是它变得较弱。同样的,当护士的持续时间比觅食的持续时间短,分工是钢筋,因为这减少护理个人的能量代谢在任务性能相对于个体觅食(无花果。S2)。

一个如果在护理代谢率较低相对于觅食,分工是钢筋。相反,分工是弱在护理期间代谢率高于在觅食。b作为他们的新陈代谢率会降低,护理人的营养水平偏离更强烈的个体觅食。图形的约定,见图。2。
支配关系强化分工,特别是主导地位与营养状况有关
资源共享通常是不平等的,但与主导地位有关18,28,29日,30.。为了研究不均匀通过优势资源共享的效果,我们分配常数主导价值观的个人仿真初始化(无花果。1 b;(3)Dominance-based共享)。在这种dominance-based摄取资源的差异,分工达到最大水平和营养水平的觅食和护理人呈双峰分布(图。4 a1)。作为个体更大优势值中获得更多的资源共享,他们迅速被固定到较低的护理而主导价值观始终饲料。中间的人主导价值观饲料或护士(图。4 a2),这是随机由随机交互,这些人在一个模拟世界中。考虑图模型场景。4缺点,分工是基于已存在的个体之间的差异(优势)。然而,同样的结果也可以获得在最初人口相同的个体,如果主导地位是由于营养水平的差异(图1 b;(4)以营养为基础的共享)。如无花果所示。4 b1,分工又达到最大水平,觅食的营养水平和护理人呈双峰分布。在仿真、营养水平的个体差异相对缓慢但是一旦存在微小的差异,一个完整的营养水平的差异正在迅速实现(图4 b2)。在无花果。S3,我们表明,分工与低营养水平甚至出现个人中获得更多的资源共享。在这个更无私的分享场景中,低水平的劳动分工开始觅食的出现,因为一些个人获得的资源返回觅食的个体,从而延迟觅食。这个结果是类似于图。3,低水平的劳动分工甚至出现觅食比护理相关的代谢率较低。

a1Dominance-based共享。分工达到最大程度的D= 1。护理和觅食的个体表现出双峰分布的营养水平。a2营养水平的个体差异在仿真时间,取决于个人的主导地位。b1以营养为基础的共享。分工又达到最大程度的D= 1,和营养水平表现出双峰分布。b2营养水平的个体差异在仿真时间。a1+b1图形的约定,见图。2。a2+b2每一行显示一个人的营养水平在前20%的模拟时间从一个代表复制模拟、彩色根据个人价值如果主导价值观是预先分配的主导地位。
分工出现不分大小
任务专业化建议,增加了群体的大小12,31日,32但这并不是明确的支持25,33。所以我们改变了集团规模调查影响分工的出现。图5表明分工出现独立的组大小。然而,在小群体中,分工的程度可能强烈依赖于细节(如是否集团大小是奇数还是偶数)。图年代4进一步阐述了这一点。
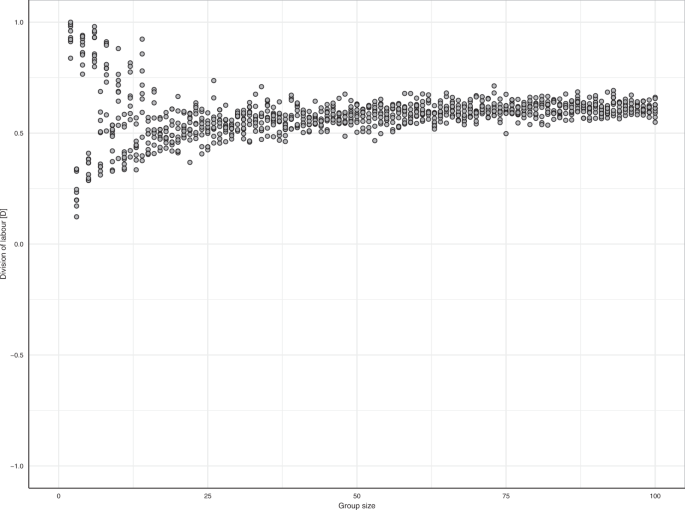
分工出现不分大小。每个点显示了一个复制的分工度量仿真每组大小(n= 10)。
讨论
我们已经表明,资源共享可能导致强烈的任务专业化和分工。个体之间的预先存在的差异可以提高分工,但他们不需要出现。我们的模型是基于两个将军和可信的假设。首先,个人营养水平下降的过程中,诱发个人出去觅食一旦变得非常低。在缺乏互动与其他个体,这导致一个活动安排,个人轮流觅食(当营养水平正在下降)和护理(补充营养水平时)。第二,如果采摘者分享一些与其他个体的收集资源,这个活动时间表就打断了由于两个反馈:个人护理可能是美联储在感知饥饿信号,从而阻碍他们的下一个觅食场;和觅食的人不能使用共享资源的充分补充营养水平,从而减少时间饥饿信号诱导他们开始觅食了。这些反馈导致任务专业化和劳动分工,由其他人因为护理个人,美联储可能继续护理而觅食个人,谁养活别人,可能会继续觅食。
大多数响应阈值模型假定个人一生都有不同的响应阈值。这些差异通常被认为是建立在生命的早期,因遗传差异9,10,11,12,13在个人发展或随机过程34,35,36。相比之下,个人在我们的模型中都有相同的阈值在整个lifetime-there特性转化在我们的模型中,但这只影响个人的瞬间的决定而不是他们的决定机制。这区别我们的模型和传统模型可以通过实验测试。如果分工出现由于个人间的差异反应阈值稳定,响应阈值模型假设,个体觅食(或护理)在一个社会群体会饲料(或护士)再次放入另一组。相比之下,如果所有人都采用相同的决策机制和分工出现通过反馈机制作用于其他参数(如个人营养水平,在我们的模型),没有这样的相关性是预期如果营养水平(或任何其他的反馈参数)在每个实验的开始扳平比分。
在某些型号的微生物专业34,35,专业化的第一步的结果从表型噪音,也就是说,从随机因素打破对称最初由诱导相同的个体小,随机差异。随后,这些差别是稳定和增强系统的特定属性或正反馈过程。我们的模型还包括表型噪声作为一个对称断路器(作为个人评估的营养水平与一些不准确),但正如图所示。2这本身并不引起分工。为此,需要资源共享(无花果。2 b)。资源共享也为微生物专业化模型中起着关键作用34,37。然而,在这些模型中,资源在全球范围内可用,从而共享的个人,而在我们的模型中个体评估他们的营养水平方面的个人财产来决定他们的任务,和资源之间共享特定的个人。
几项研究已经提出,劳动分工在社会性昆虫觅食和护士与营养相关的差异16,17,18,19,20.,21,22,38,39,40。在我们的模型中,劳动分工导致情况营养水平的护士高于觅食,和分工时提高代谢率高于觅食时,护理时代谢率。然而,不需要预先存在的营养差异的出现了工人分工至关重要的原料是资源共享。资源可以在社会性昆虫,例如,通过trophallactic口对口交流转移24。然而,对于分工的出现不是必需的,无私地共享资源。觅食,例如,存款的一部分资源巢(注定要喂小鸡在稍后的阶段),他们被其他个体。
我们这里工作分工的定义基于单个一致性和个体之间的变化在任务中选择一组的成员1。这个定义允许自发分工出现的可能性从调控机制尚未进化选择的分工,但用于其他目的。另外其他分工的定义强调个人之间的相关性选择合作3。等的进化模型中合作分工,商品的shareability /好处对劳动分工的发展至关重要37,41。我们的模型不包括进化,作为机制自我组织分工的出现(hunger-induced觅食和资源共享)认为是事先存在,不改变在我们的模拟。然而,这样non-evolved分工可以具有重要的进化意义,因为它可以后续适应性进化的衬底。这是优雅的一个实验合作机器人所示42,更容易适应分工演化在预先存在的行为差异的存在。同样,一些进化理论在社会分工的进化昆虫表明最初的步骤是基于已有的机制,监管行为单独的祖先43,44,45。
我们的模型进一步突出优势互动和特定于任务的重要性不同的营养水平很强的非生殖分工的出现。优势和营养水平也发挥重要作用在社会性昆虫的生殖能力的决心28,29日,30.,46,47而在社会蜘蛛48,因此在生殖分工。例如,在纸黄蜂、增殖和助手的角色很可能由营养49还表示,贮藏蛋白质的微分表达式育种家和帮手50,51。此外,在群居昆虫,种姓通常由nutrition-dependent发育开关52在群居昆虫,种姓差异表达基因编码存储蛋白质卵黄蛋白原40,53,54,55,56。因此,营养水平的差异,出现在我们的模型中由于在资源共享优势交互也可能为真社会性的发展起着重要的作用。
响应阈值模型和实证研究12,31日,32(尽管不是全部25,33)表明,任务专业化和劳动分工在大群最为明显。在我们的模型中,分工出现即使在非常小的群体。事实上,分工可以更强(如两组)在小群体中比在更大的群体。同样,实验表明,分工出现配对关联的蚂蚁18,57,一些种类的鸟类的性别表现出很强的繁殖的雌性之间的分工和觅食的男性58,59,60。因此,劳动分工的强度并不一定取决于组织规模也是社会交往的细节,如个体的觅食者共享资源的数量,被认为是至关重要的。
最后,资源共享的机制的出现分工超越动物组。例如,有些蓝藻细胞是专门在殖民地获得不同resources-carbon或为氮气61年。资源共享可能发挥作用在细胞的出现和监管分工中主要进化过渡到多细胞生物62年,63年。
总的来说,我们的模型表明,自我组织分工出现相同的个体之间通过资源共享。鉴于生物无所不在的资源共享系统的实例,在群居的动物,群居昆虫,人类的狩猎社会,或在多细胞organisms-our模型显示一种机制,可以解释分工的出现在很大范围内的生物。
方法
该模型
我们开发了一个基于单独的仿真模型在连续时间。每个模拟代表一群N个人(N= 100,除非另有规定)。个人饲料获取资源或执行其他任务,我们称作为护理。每一个仿真时间T时间的步骤(T= 10000)。
营养水平的个体
个人拥有一个内部状态变量n这反映了他们的营养水平和范围从0到n马克斯(n马克斯= 100)。在模拟的开始,所有的初始化的人与一个相同营养水平n初始化(n初始化= 50)。因此,模拟从相同的个体。随着时间的推移,个人的营养水平降低代谢率米为当觅食,米努尔当护理(米为= 1.0,米努尔= 1.0,除非另有规定)。
任务的选择
饲料或护士个人的选择取决于个人的营养水平。我们假设个体成为更容易饲料营养水平下降。平均而言,个人开始觅食当营养水平达到临界阈值\μ(\ \)(\ \(μ= 50 \))。然而,我们假设并不完全准确评估个体的营养水平:感知到的营养水平是来自正态分布在真正的营养水平,标准偏差σ\ (\ \)(\(σ= 1 \ \))。从觅食之旅回来后,有一个固定的时间t为时间的步骤(t为= 5),个体之间的选择再次觅食或切换到护理。选择取决于个人的感知营养水平与觅食的阈值:个人将再次饲料如果认为营养水平低于阈值,否则,它会切换到护理。资源可以通过资源共享来获得个人护理,其中包括先前觅食个人,填充并切换回护理的营养水平。个人因此没有预期执行护理或觅食任务和所有个人原则上可以分享和获得资源。如果前护理个人不是美联储认为营养水平下降低于阈值,然后它开始觅食。如果美联储护理个人觅食,然后处理食品在固定的时间t努尔时间的步骤(t努尔= 5),在此期间无法收到任何进一步的资源。食品加工后,个人护理评估他们的营养水平和决定是否继续护理或开始觅食(无花果。S5)。
营养转移
觅食者获得R资源而觅食(R= 10)。我们实现了四个不同的模型场景的资源共享。(1)没有共享:从觅食之旅回来后,觅食的人让他们获得的所有资源。(2)平等共享:从觅食之旅回来后,觅食个人均匀地共享资源我个人护理。资源的数量获得个人从事的相互作用是如此\ (R / (1 + i) \)因为一个觅食个人与之交互我个人护理。我们假设我= 1,但放松这个假设的补充材料(无花果。S4)。(3)Dominance-based共享:个人分配一个主导价值的初始化模拟,从一个均匀分布的随机抽样从0到1。期间获得的资源数量的个体相互作用依赖于他们的优势价值,这样比少占主导地位的个人获得更多的资源占主导地位的个人。个人获得的资源的数量k计算使用将softmax函数
在哪里dk焦的主导价值个人吗k,dl其他的主导价值吗我个人和年代为将softmax函数是一个参数。如果年代= 0,该模型降低了平等共享的情况下,如果年代< 0,个人地位最低的值接收过多的资源,如果年代> 0,个人最高的主导价值观收到过多的资源。对于模拟的优势效应,我们使用年代= 1,导致最主要的个人获得略超过他们的预期基于他们的相对优势。(4)以营养为基础的共享:个体的主导地位是由其营养水平,相对于最大的营养水平。dk现在是由n/n马克斯。否则,交互与场景3中进行,用情商。1)来确定个体之间的资源分配。再一次,我们使用年代与其他的值= 1(模拟年代,见图。S3)。作为主导地位又决定了个体之间的资源分配,个人有更高的营养水平获得更多的资源比个人营养水平较低。
觅食时个人回报觅食之旅,但没有护理的人可以获得资源可用,觅食的个人消费的所有资源本身,独立于分享的场景。
量化的劳动分工
我们分工定义为单个的一致性,但在个人可变性在任务中选择一组的成员1。在个体一致性和个体之间的变化在单一指标很难量化64年,我们使用四个不同的指标量化的程度分工出现的模拟25,31日,64年。这四个指标不同的强调劳动分工的两个定义属性,但如无花果所示。S1他们为主要的模型场景产生相似的结果。在主要的手稿,因此我们只报告度量的结果D提出的Duarte et al。25,这是容易理解和计算。计算D,一个认为所有任务的选择情况(见图5当任务选择的情况发生),个人必须决定是否再次执行前面的任务或切换到其他任务。它被定义为:
在哪里\(酒吧\ {q} \)是个人选择的比例的情况下再次执行前面的任务(平均超过所有个人和选择的情况下),在吗\ ({p} _ {1} \)和\ ({p} _ {2} = 1 - p \)是每个两个任务的相对频率选择(觅食和护理)(平均超过所有个人和选择的情况下)。这个词\ ({p} _ {1} ^ {2} + {p} _ {2} ^ {2} \)在分母上的情商。2)对应的概率随机选择再次执行前面的任务。因此,D是正的,如果酒吧\ (\ {q} > {p} _ {1} ^ {2} + {p} _ {2} ^ {2} \),也就是说,如果个人有一个高于随机趋势再次执行前面的任务。作为\(酒吧\ {q} \)范围在0和1之间,D−1(当之间的范围酒吧\ (\ {q} = 0 \)),\ (2 p {} _ {1} / {p} _ {2} ({p} _ {1} ^ {2} + {p} _ {2} ^ {2}) \)(当酒吧\ (\ {q} = 1 \))。大约在我们的模型中,两个任务都执行在任何场景下都以同样的频率(\ ({p} _{1} \, \大约\ p {} _ {2} \))。因此,D不等−1(表示任务之间交替),+ 1(表示完整任务专业化)25。我们计算分工指标在过去10%的模拟时间避免测量初始化效果。
模型分析
c++实现的模型,用g++ 9.3.0编译。模型结果分析,在R 4.1.0形象化65年使用包ggplot266年,gridExtra67年,cowplot68年和MetBrewer69年。
报告总结
进一步研究信息设计是可用的自然投资组合报告总结与这篇文章有关。
数据可用性
在本研究可在所有数据生成https://doi.org/10.34894/WD4EJZ。
代码的可用性
模拟代码和数据分析脚本可用https://doi.org/10.34894/WD4EJZ。
引用
乌尔里希,Y。,Saragosti, J., Tokita, C. K., Tarnita, C. E. & Kronauer, D. J. C. Fitness benefits and emergent division of labour at the onset of group living.自然560年,635 - 638 (2018)。
杜阿尔特,一个。,Weissing, F. J., Pen, I. & Keller, L. An evolutionary perspective on self-organized division of labor in social insects.为启生态。另一个星球。系统。42,91 - 110 (2011)。
西方,s . a &库珀g . a .微生物分工:进化的观点。启Microbiol Nat。14,716 - 723 (2016)。
奥斯特,g . f . & Wilson e . O。种姓制度和生态社会昆虫。(普林斯顿大学出版社,1978)。
阿诺德·k·E。,Owens, I. P. F. & Goldizen, A. W. Division of labour within cooperatively breeding groups.Behav142年,1577 - 1590 (2005)。
Bruintjes, r & Taborsky m .尺度依赖的任务分工合作丽鱼科鱼,以应对实验需求的变化。动画。Behav。81年,387 - 394 (2011)。
Bergmuller, r & Taborsky m .适应性行为综合症由于战略细分专业化。BMC生态。712 (2007)。
夏勒,g B。塞伦盖蒂狮子:捕食关系的研究。(芝加哥大学出版社,2009)。
拿E。,Theraulaz, G. & Deneubourg, J.-L. Quantitative study of the fixed threshold model for the regulation of division of labour in insect societies.Proc,杂志。科学。263年,1565 - 1569 (1996)。
拿大肠固定响应阈值和在昆虫社会劳动分工的规定。公牛。数学。医学杂志。60,753 - 807 (1998)。
格雷厄姆,S。,Myerscough, M. R., Jones, J. C. & Oldroyd, B. P. Modelling the role of intracolonial genetic diversity on regulation of brood temperature in honey bee (Apis mellifera L.) colonies.昆虫Soc。53,226 - 232 (2006)。
Jeanson, R。,Fewell, J. H., Gorelick, R. & Bertram, S. M. Emergence of increased division of labor as a function of group size.Behav。生态。Sociobiol。62年,289 - 298 (2007)。
戈夫,R。,Hayworth, M., Chhetri, M. & Rueppell, O. Division of labour and social insect colony performance in relation to task and mating number under two alternative response threshold models.昆虫。Soc。56,319 - 331 (2009)。
乌尔里希,y . et al .响应阈值无法解释经验的分工模式在社会昆虫。公共科学图书馆杂志。19e3001269 (2021)。
Jeanson r & Weidenmuller a个人间变异性在社会性昆虫——近因和最终的后果。医学杂志。牧师。89年,671 - 687 (2014)。
托斯a . l .罗宾逊& g . e .工人在蜜蜂营养和分工。动画。Behav。69年,427 - 435 (2005)。
史密斯,c . r . et al .营养不对称有关无女王的蚂蚁的分工。《公共科学图书馆•综合》6e24011 (2011)。
Bernadou, a . et al .压力和早期经验基础优势地位和分工在克隆昆虫。Proc。r . Soc。B285年,20181468 (2018)。
Bernadou,。,Hoffacker, E., Pable, J. & Heinze, J. Lipid content influences division of labour in a clonal ant.j . Exp。杂志。223年杰布。219年238(2020).
Dussutour,。,Poissonnier, L.-A., Buhl, J. & Simpson, S. J. Resistance to nutritional stress in ants: when being fat is advantageous.j . Exp。杂志。219年,824 - 833 (2016)。
布兰查德,g B。、Orledge g M。,Reynolds, S. E. & Franks, N. R. Division of labour and seasonality in the ant Leptothorax albipennis: worker corpulence and its influence on behaviour.动画。Behav。59,723 - 738 (2000)。
托斯,a . L。,Kantarovich, S., Meisel, A. F. & Robinson, G. E. Nutritional status influences socially regulated foraging ontogeny in honey bees.j . Exp。杂志。208年,4641 - 4649 (2005)。
卡特,g . g . &威尔金森g s吸血蝙蝠的食品分享:相互帮助预测捐款超过亲缘或骚扰。Proc。r . Soc。B280年,20122573 (2013)。
Meurville显示,& LeBoeuf AdriaC。交哺:蚁群中功能和社会的进化液交换(膜翅目:蚁科)。Myrmecol N。31日行(2021)。
杜阿尔特,一个。,我的钢笔。,Keller, L. & Weissing, F. J. Evolution of self-organized division of labor in a response threshold model.Behav。生态。Sociobiol。66年,947 - 957 (2012)。
摩尔,K。,Federle, W. & Roces, F. The energetics of running stability: costs of transport in grass-cutting ants depend on fragment shape.j . Exp。杂志。215年,161 - 168 (2012)。
奥斯特瓦尔德,M . M。福克斯,t . P。,Harrison, J. F. & Fewell, J. H. Social consequences of energetically costly nest construction in a facultatively social bee.Proc。r . Soc。B288年,20210033 (2021)。rspb.2021.0033。
莫利纳,y & O ' donnell美国发展dominance-nutrition假说的测试:将成年喂食,侵略,paperwasp繁殖潜能Mischocyttarusmastigophorus。Ethol。生态。另一个星球。20.,125 - 139 (2008)。
达。菲奥卡,并于当年k . et al .生殖生理学对应于成人营养和任务绩效在新热带区的胡蜂:测试dominance-nutrition假说预测。Behav。生态。Sociobiol。74年114 (2020)。
Wcislo, w . t . &冈萨雷斯,v . h .社会和生态环境的交哺兼性社会汗水蜜蜂,Megalopta genalis和m . ecuadoria(膜翅目昆虫,Halictidae)。昆虫Soc。53,220 - 225 (2006)。
Gautrais, J。,Theraulaz, G., Deneubourg, J.-L. & Anderson, C. Emergent polyethism as a consequence of increased colony size in insect societies.j理论的。医学杂志。215年,363 - 373 (2002)。
Ferguson-Gow, H。萨姆纳,S。,Bourke, A. F. G. & Jones, K. E. Colony size predicts division of labour in attine ants.Proc。r . Soc。B281年,20141411 (2014)。
多恩豪斯,一个。,Holley, J.-A. & Franks, N. R. Larger colonies do not have more specialized workers in the ant Temnothorax albipennis.Behav。生态。20.,922 - 929 (2009)。
阿克曼,m . et al .自我毁灭的合作由表型噪音。自然454年,987 - 990 (2008)。
Dubnau d & Losick, r .双稳性细菌。摩尔。Microbiol61年,564 - 572 (2006)。
Honegger k & de Bivort b .特性转化、个性和行为。咕咕叫。医学杂志。28R8-R12 (2018)。
Schiessl, k . t . et al .个人与group-optimality生产细菌分泌的化合物。进化73年,675 - 688 (2019)。
Elsner D。,Hartfelder, K. & Korb, J. Molecular underpinnings of division of labour among workers in a socially complex termite.科学。代表。1118269 (2021)。
Kohlmeier, P。,Feldmeyer, B. & Foitzik, S. Vitellogenin-like A–associated shifts in social cue responsiveness regulate behavioral task specialization in an ant.公共科学图书馆杂志。16e2005747 (2018)。
Morandin C。,Hietala, A. & Helanterä, H. Vitellogenin and vitellogenin-like gene expression patterns in relation to caste and task in the ant Formica fusca.昆虫Soc。66年,519 - 531 (2019)。
库珀·g·a . &西方,s . a的劳动分工和极端的专业化的进化。Nat,生态。另一个星球。2,1161 - 1167 (2018)。
费,E。,Turgut, A. E., Duéñez-Guzmán, E., Dorigo, M. & Wenseleers, T. Evolution of Self-Organized Task Specialization in Robot Swarms.公共科学图书馆第一版。医学杂志。11e1004273 (2015)。
West-Eberhard, M.J.黄蜂社会的微观研究的发展和演化。在paper-wasps的自然历史和演化(eds。Turillazzi, s & West-Eberhard m . j .) 290 - 317(牛津大学出版社,1996年)。
West-Eberhard, m . j .灵活的策略和社会进化。在动物社会:理论和事实(eds。ItōY。,Brown, J. L. & Kikkawa, J.) 35–51 (Japan Scientific Societies Press, 1987).
Amdam教授,g . V。,Csondes, A., Fondrk, M. K. & Page, R. E. Complex social behaviour derived from maternal reproductive traits.自然439年,76 - 78 (2006)。
克里,j . U。梵天,。,Chavan, S. K. & Gadagkar, R. Nutrition induced direct fitness for workers in a primitively eusocial wasp.昆虫Soc。68年,319 - 325 (2021)。
O ' donnell et al。成人营养和生殖生理学:稳定同位素分析在群居胡蜂(Mischocyttarus mastigophorus,膜翅目:胡蜂科)。Behav。生态。Sociobiol。72年86 (2018)。
所罗门,M。,Mayntz, D. & Lubin, Y. Colony nutrition skews reproduction in a social spider.Behav。生态。19,605 - 611 (2008)。
亨特,j . h & Amdam教授g . v . Bivoltinism作为前期胡蜂属Polistes真社会性。科学308年,264 - 267 (2005)。
亨特,j . H。,Buck, N. A. & Wheeler, D. E. Storage proteins in vespid wasps: characterization, developmental pattern, and occurrence in adults.j .昆虫杂志。49,785 - 794 (2003)。
亨特,j . h . et al .微分基因表达和蛋白质丰度表明个体发生的倾向种姓在最初地群居的黄蜂。《公共科学图书馆•综合》5e10674 (2010)。
史密斯,c R。,托斯,a . L。,Suarez, A. V. & Robinson, G. E. Genetic and genomic analyses of the division of labour in insect societies.Nat,启麝猫。9,735 - 748 (2008)。
萨姆纳,S。,Pereboom, J. J. M. & Jordan, W. C. Differential gene expression and phenotypic plasticity in behavioural castes of the primitively eusocial wasp,Polistes黄花。Proc。r . Soc。B273年19-26 (2006)。
格拉夫,J。,Jemielity, S., Parker, J. D., Parker, K. M. & Keller, L. Differential gene expression between adult queens and workers in the ant Lasius niger.摩尔。生态。16,675 - 683 (2007)。
纳尔逊,c . M。Ihle, k . E。,Fondrk, M. K., Page, R. E. & Amdam, G. V. The gene vitellogenin has multiple coordinating effects on social organization.公共科学图书馆杂志。5约(2007)。
电晕,m . et al .卵黄蛋白原接受subfunctionalization收购种姓和行为中的特定表达收割机ant Pogonomyrmex barbatus。公共科学图书馆麝猫9e1003730 (2013)。
周期,j . h &页面,r·e·Jr分工的出现迫使协会通常孤独的蚂蚁皇后。Evolut。生态。Res。1,537 - 548 (1999)。
Kalina j .巢入侵者巢国防和觅食行为的黑白盔犀鸟Bycanistes subcylindricus。宜必思131年,567 - 571 (1988)。
Heinsohn, r & Legge reverse-dichromatic的繁殖生物学,合作鹦鹉Eclectus roratus。j .黑旋风。259年,197 - 208 (2003)。
Zarybnicka, m & Vojar j .男性供应对父母行为的影响女性的北方猫头鹰Aegolius funereus。黑旋风。钉。5236 (2013)。
弗洛雷斯,e . &并没有区分函数通过丝状蓝藻的细胞分化。启Microbiol Nat。839-50 (2010)。
梅纳德史密斯,j . & Szathmary E。在进化过程中主要的过渡。(w·h·弗里曼,1995)。
西方,美国。,Fisher, R. M., Gardner, A. & Kiers, E. T. Major evolutionary transitions in individuality.Proc。《科学。112年,10112 - 10119 (2015)。
Gorelick, R。,Bertram, S. M., Killeen, P. R. & Fewell, J. H. Normalized mutual entropy in biology: quantifying division of labor.点。博物学家164年,677 - 682 (2004)。
R核心团队。接待员:统计计算的语言和环境。(R统计计算的基础,2021)。
韦翰,H。ggplot2:优雅的图形进行数据分析。纽约(斯普林格出版社,2016)。
Auguie B。“网格”图形gridExtra:杂项功能。(https://CRAN.R-project.org/package=gridExtra,2017)。
Wilke, c . O。cowplot:流线型的情节主题和情节注释为“ggplot2”。(https://CRAN.R-project.org/package=cowplot,2019)。
米尔斯,b R。MetBrewer:调色板灵感来自在大都会艺术博物馆工作。(https://CRAN.R-project.org/package=MetBrewer,2021)。
确认
我们感谢汉诺Hildenbrandt帮助我们调试模型,提高仿真效率。我们感谢格罗宁根大学的信息技术中心提供访问游隼高性能计算集群。J.J.K.被一个自适应支持格兰特格罗宁根大学的生活。他还是被DFG格兰特没有支持。BE6684/1−1(研究单位“很长时间”,2218)。D.E.被DFG格兰特没有支持。朱迪思科博KO1895/19−1()和NWO批准号823.01.006 (F.J.W.)。F.J.W.承认资金从欧洲研究委员会(先进伦理委员会批准号789240)。
作者信息
作者和联系
贡献
范本:J.J.K.,T.J.,学士,D.E.,B。H.K., F.J.W.; Implementation: T.J., J.J.K.; Model analysis: J.J.K., D.E.; Writing: J.J.K., T.J., A.B., D.E., B.H.K., F.J.W.
相应的作者
道德声明
相互竞争的利益
作者宣称没有利益冲突。
同行评审
同行审查的信息
自然通讯感谢匿名评论者对他们的贡献的同行评审工作。同行审查报告是可用的。
额外的信息
出版商的注意施普林格自然保持中立在发表关于司法主权地图和所属机构。
权利和权限
开放获取本文是基于知识共享署名4.0国际许可,允许使用、共享、适应、分布和繁殖在任何媒介或格式,只要你给予适当的信贷原始作者(年代)和来源,提供一个链接到Creative Commons许可,并指出如果变化。本文中的图片或其他第三方材料都包含在本文的创作共用许可,除非另有说明在一个信用额度的材料。如果材料不包括在本文的创作共用许可证和用途是不允许按法定规定或超过允许的使用,您将需要获得直接从版权所有者的许可。查看本许可证的副本,访问http://creativecommons.org/licenses/by/4.0/。
关于这篇文章

引用这篇文章
罗斯·j。简森,T。,Bernadou,。et al。资源共享是充分的劳动分工的出现。Nat Commun137232 (2022)。https://doi.org/10.1038/s41467 - 022 - 35038 - 2
收到了:
接受:
发表:
DOI:https://doi.org/10.1038/s41467 - 022 - 35038 - 2
