








文摘gydF4y2Ba
溶解氧(OgydF4y2Ba2gydF4y2Ba)是至关重要的对于大多数海洋生态系统,推动生物的呼吸作用和促进碳和养分的循环。氧气测量被解释为表明海洋缺氧的区域(ODZs)正在扩大在全球变暖gydF4y2Ba1gydF4y2Ba,gydF4y2Ba2gydF4y2Ba。然而,模型提供了一个清楚的照片未来ODZ变化在短期和长期的gydF4y2Ba3gydF4y2Ba,gydF4y2Ba4gydF4y2Ba,gydF4y2Ba5gydF4y2Ba,gydF4y2Ba6gydF4y2Ba。古气候记录可以帮助探索的可能范围ODZ warmer-than-modern时期的变化。在这里我们使用foraminifera-bound氮(N)同位素表明潜脱氮在北太平洋东部的热带大大减少在中新世中期气候适宜期(MMCO)和始新世早期气候适宜期(EECO)。因为脱氮是局限于缺氧的水域,我们的研究结果表明,在这两个新生代时期的持续温暖,ODZs简约,不扩大。ODZ收缩可能出现下降upwelling-fuelled热带太平洋生物生产力,这将减少需氧量在地下。另外,滋补深水通风的南大洋可能削弱海洋的“生物碳泵”,这将增加了深海氧。机制在起作用就会决定是否ODZ收缩发生在与全球变暖或花了几百年或几千年发展。因此,虽然我们的新生代的结果不一定适用于短期的未来,他们可能暗示全球变暖可能最终导致ODZ收缩。gydF4y2Ba
主要gydF4y2Ba
观测表明,氧气浓度在沿海和公海水域已减少在过去的五十年,大幅扩大海洋ODZs的体积gydF4y2Ba1gydF4y2Ba,gydF4y2Ba2gydF4y2Ba。全球脱氧趋势归因于减少氧气溶解度和增强的上层海洋分层预计从全球变暖,全球氧浓度预测,在未来几十年将继续减少,几个世纪以来,影响海洋生态系统gydF4y2Ba7gydF4y2Ba。然而,有争议的未来ODZs特别是与预测的扩张和收缩gydF4y2Ba3gydF4y2Ba,gydF4y2Ba4gydF4y2Ba,gydF4y2Ba5gydF4y2Ba,gydF4y2Ba6gydF4y2Ba。gydF4y2Ba
地球的气候演变响应构造和轨道迫使新生代,过去的6600万年里(最高产量研究)。总的来说,大气有限公司gydF4y2Ba2gydF4y2Ba浓度下降和全球气候已经冷却gydF4y2Ba8gydF4y2Ba。在此背景下新生代的冷却,有长时间的温暖气候的两个不同的时期:MMCO,大约6°C比今天暖和,EECO,约13°C比今天暖和gydF4y2Ba9gydF4y2Ba。MMCO和EECO的特点是高有限公司gydF4y2Ba2gydF4y2Ba(分别为550 ppm左右和1650 ppm)和减少或没有大陆冰gydF4y2Ba10gydF4y2Ba,gydF4y2Ba11gydF4y2Ba,gydF4y2Ba12gydF4y2Ba。这两个气候最佳状态提供了一个机会来调查海洋氧气浓度长时间温暖的回应gydF4y2Ba13gydF4y2Ba。gydF4y2Ba
海洋中N同位素的比率是对缺氧敏感,或“suboxia”(溶解OgydF4y2Ba2gydF4y2Ba浓度低于约5µmol公斤gydF4y2Ba−1gydF4y2Ba基于标准的测量)。硝酸细菌减少到NgydF4y2Ba2gydF4y2Ba(“脱”)需要suboxia和歧视gydF4y2Ba15gydF4y2Ban .因此,脱氮的ODZs海水柱引起的高程gydF4y2Ba15gydF4y2BaN /gydF4y2Ba14gydF4y2BaN比率,或δgydF4y2Ba15gydF4y2BaODZs N,硝酸的地区gydF4y2Ba14gydF4y2Ba,也增加了硝酸盐δgydF4y2Ba15gydF4y2Ba全球海洋的NgydF4y2Ba15gydF4y2Ba(δgydF4y2Ba15gydF4y2BaN = ((gydF4y2Ba15gydF4y2BaN /gydF4y2Ba14gydF4y2BaN)gydF4y2Ba样本gydF4y2Ba/ (gydF4y2Ba15gydF4y2BaN /gydF4y2Ba14gydF4y2BaN)gydF4y2BaatmN2gydF4y2Ba−1)×1000‰)。ODZ脱氮的同位素信号是并入新成立的生物在海洋表面,在深海沉积物中积累的副产品。因此,N同位素为重建提供一个工具过去ODZ-hosted脱氮的变化gydF4y2Ba16gydF4y2Ba。的有机质和矿物矩阵内被困planktic有孔虫壳(foraminifer-bound有机物)已经被开发作为存档的N同位素海洋表面生产力的历史gydF4y2Ba17gydF4y2Ba。Foraminifer-bound有机物被孤立于矿物沉积环境的矩阵,保留原始不变的N同位素信号甚至超过数百万年gydF4y2Ba18gydF4y2Ba。在硝酸完成消费地区,foraminifer-boundδgydF4y2Ba15gydF4y2BaN记录δgydF4y2Ba15gydF4y2BaN硝酸的底层密度跃层gydF4y2Ba19gydF4y2Ba。硝酸密度跃层δgydF4y2Ba15gydF4y2BaN熊区域信号潜反硝化和NgydF4y2Ba2gydF4y2Ba固定,可用生物的主要来源(即“固定”)N的海洋gydF4y2Ba20.gydF4y2Ba。硝酸密度跃层δgydF4y2Ba15gydF4y2BaN也将随δgydF4y2Ba15gydF4y2BaN硝酸的意思是海洋。意味着海洋硝酸δgydF4y2Ba15gydF4y2BaN,反过来,在很大程度上是由脱氮的相对比例控制发生在水体和沉积物gydF4y2Ba15gydF4y2Ba。gydF4y2Ba
N同位素和温度重建gydF4y2Ba
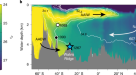
在这项研究中,我们测量δgydF4y2Ba15gydF4y2BaN foraminifer-bound有机物(FB-δgydF4y2Ba15gydF4y2BaN)调查新生代海洋N循环的变化。我们现在新菌种FB-δgydF4y2Ba15gydF4y2BaN中新世以来的数据,以及genus-specific测量在中新世早期,渐新世和晚始新世时代从两个沉积物,一个位于太平洋(872年ODP网站),另一个在大西洋(DSDP站点516)(图。gydF4y2Ba1gydF4y2Ba)。这些数据编译之前发表mixed-taxa和genus-specific FB-δgydF4y2Ba15gydF4y2BaN在太平洋ODP数据从始新世和古新世时代网站1209年和1263年大西洋ODP站点,分别(无花果。gydF4y2Ba1gydF4y2Ba),合并后的数据集覆盖的新生代(无花果。gydF4y2Ba2摄氏度gydF4y2Ba和扩展数据图。gydF4y2Ba1gydF4y2Ba)。gydF4y2Ba

地质构造变化驱动的站点位置显示符号大小(见图例)。圆圈表示核心装满FB-δgydF4y2Ba15gydF4y2BaN数据,从这项研究中(872/516)和清晨里等。gydF4y2Ba18gydF4y2Ba(1209/1263),与有孔虫δ和开放的圆圈显示核心gydF4y2Ba18gydF4y2BaO数据讨论的文本gydF4y2Ba37gydF4y2Ba,gydF4y2Ba41gydF4y2Ba(引用)。解散了啊gydF4y2Ba2gydF4y2Ba浓度(μmol公斤gydF4y2Ba−1gydF4y2Ba)所示的颜色。gydF4y2Ba

一个gydF4y2Ba海温资料,编制新的和以前发表的基于特克斯gydF4y2Ba86年gydF4y2Bapalaeothermometer分为高纬度地区(高于50°N / S)和低/中纬度地区(0-40°N / S),与黄土平滑(0.02因素)和90%置信区间gydF4y2Ba方法gydF4y2Ba和补充数据gydF4y2Ba2gydF4y2Ba)。特克斯gydF4y2Ba86年gydF4y2Ba温度的估计是基于BAYSPAR校准(见gydF4y2Ba方法gydF4y2Ba)。灰色的线显示了底栖有孔虫氧同位素比率(δgydF4y2Ba18gydF4y2BaO)编译Westerhold et al。gydF4y2Ba8gydF4y2Ba。gydF4y2BabgydF4y2Ba,ΔδgydF4y2Ba15gydF4y2BaNgydF4y2Ba便士gydF4y2Ba是FB-δgydF4y2Ba15gydF4y2BaN ODP网站872太平洋和大西洋之间的区别DSDP网站516(黑线)或太平洋ODP网站1209至1263年大西洋ODP网站(灰色线)。ΔSSTgydF4y2BaL-HgydF4y2Ba(红线)海温之间的区别是低/中纬度和高纬度地区(从gydF4y2Ba一个gydF4y2Ba;ΔSST传播90%置信区间gydF4y2BaL-HgydF4y2Ba扩展数据图所示。gydF4y2Ba4 bgydF4y2Ba)。gydF4y2BacgydF4y2BaFB-δ,平均的种特异性gydF4y2Ba15gydF4y2BaN从ODP站点872(绿色)和DSDP网站516(蓝色),连同mixed-taxa FB-δgydF4y2Ba15gydF4y2BaN从ODP站点1209(浅绿色)和ODP网站1263(浅蓝色),两个尺寸的计算的平均分数gydF4y2Ba18gydF4y2Ba。三角形和圆形的不同颜色显示个人种专一性或genus-specific FB-δgydF4y2Ba15gydF4y2Ba分别在网站网站872/1209和516/1263 N(见扩展数据图。gydF4y2Ba1gydF4y2Ba详情)。亮粉红色酒吧的最初迹象表明全球变暖到EECO MMCO,温暖和暗条显示主要的间隔,基于底栖有孔虫δgydF4y2Ba18gydF4y2BaO (gydF4y2Ba一个gydF4y2Ba)。比如,更新世;Pli,上新世;朋友,古新世。gydF4y2Ba
此外,我们已经生成了特克斯gydF4y2Ba86年gydF4y2Ba的gydF4y2Ba21gydF4y2Ba海表面温度(SST)编译,包括新的测量以及以前公布的数据(无花果。gydF4y2Ba2gydF4y2Ba无花果和扩展数据。gydF4y2Ba2gydF4y2Ba和gydF4y2Ba3gydF4y2Ba)。编译显示上层海洋的热演化在低/中、高纬度地区。对海温编译允许我们重建low-to-high-latitude SST梯度(ΔSSTgydF4y2BaL-HgydF4y2Ba使用一个温度代理(图)。gydF4y2Ba2 bgydF4y2Ba无花果和扩展数据。gydF4y2Ba2gydF4y2Ba- - - - - -gydF4y2Ba4gydF4y2Ba)。ΔSSTgydF4y2BaL-HgydF4y2Ba可能是一个特别敏感的过去气候变化的指标,并提供在大气和海洋环流的约束。gydF4y2Ba
新的数据显示,大西洋和太平洋FB-δgydF4y2Ba15gydF4y2BaN值升高对目前在中新世早期但大幅减少两个盆地从19岁到1600万年前(Ma),和一个更大的下降在太平洋(约11‰)比大西洋(大约6‰)(无花果。gydF4y2Ba2摄氏度gydF4y2Ba)。FB-δgydF4y2Ba15gydF4y2BaN在MMCO达到最低两个站点。FB-δgydF4y2Ba15gydF4y2BaN然后增加在随后的冷却阶段(马马从14到10),称为中间中新世气候转变,再次与更大的改变在太平洋大西洋。最后,在中新世晚期(马马从10到5),记录分离:太平洋记录显示FB-δ强劲gydF4y2Ba15gydF4y2BaN增加之后,马3的下降,而大西洋记录首先显示一个小降低然后保持稳定,直到现在。gydF4y2Ba
编译后的数据显示为MMCO和EECO收敛行为。气候最适条件都是太平洋FB-δ低的特征gydF4y2Ba15gydF4y2BaFB-δN和减少gydF4y2Ba15gydF4y2BaN(Δδ太平洋和大西洋之间的区别gydF4y2Ba15gydF4y2BaNgydF4y2Ba便士gydF4y2Ba;无花果。gydF4y2Ba2 b, cgydF4y2Ba)。此外,气候最适条件都与强烈的高纬度地区的海洋变暖,减少了子午SST梯度(图。gydF4y2Ba2 a, bgydF4y2Ba)。ΔδgydF4y2Ba15gydF4y2BaNgydF4y2Ba便士gydF4y2Ba和ΔSSTgydF4y2BaL-HgydF4y2Ba相关不仅在升温阶段,而且更广泛地通过新生代(无花果。gydF4y2Ba2 bgydF4y2Ba)。gydF4y2Ba
减少suboxia温暖的时期gydF4y2Ba
δ的耦合的下降gydF4y2Ba15gydF4y2BaN和ΔδgydF4y2Ba15gydF4y2BaNgydF4y2Ba便士gydF4y2Ba在EECO MMCO最好解释为减少潜脱氮,现在大多发生在热带太平洋东部ODZs(无花果。gydF4y2Ba1gydF4y2Ba)。潜脱氮的下降,缺乏类似的底栖生物脱氮的下降,降低了δgydF4y2Ba15gydF4y2BaN硝酸的意思是海洋gydF4y2Ba15gydF4y2Ba因此,δgydF4y2Ba15gydF4y2BaN在两个记录。同时,如果suboxia和潜脱氮持续一直集中在太平洋,潜脱氮的下降将减少热带太平洋硝酸盐δ的高程gydF4y2Ba15gydF4y2BaN相对于其他海洋,降低ΔδgydF4y2Ba15gydF4y2BaNgydF4y2Ba便士gydF4y2Ba观察到的。因此,与普遍的预期gydF4y2Ba22gydF4y2Ba,我们FB-δgydF4y2Ba15gydF4y2BaN数据显示减少,而不是一个提高,ODZs最热的时间间隔期间的新生代。gydF4y2Ba
两个FB-δgydF4y2Ba15gydF4y2BaN和ΔδgydF4y2Ba15gydF4y2BaNgydF4y2Ba便士gydF4y2Ba在EECO开始大幅下跌。亚洲印度碰撞的影响上流出的特提斯海已被建议作为重要的变化,通过其对海洋的密度跃层的温度的影响gydF4y2Ba18gydF4y2Ba。类似的机制可能为重建提供反硝化MMCO开始下降,当proto-Mediterranean接受更大的隔离从印度洋gydF4y2Ba23gydF4y2Ba。然而,这种Tethys-outflow-focused解释并不能解释后续增加太平洋潜脱氮增加Δδ所示gydF4y2Ba15gydF4y2BaNgydF4y2Ba便士gydF4y2Ba后每一个暖期(无花果。gydF4y2Ba2 bgydF4y2Ba)。gydF4y2Ba
其他值得注意的构造变化大致重叠MMCO结束。其中包括扩大15岁的德雷克海峡马在10-Myr时间的缩小gydF4y2Ba24gydF4y2Ba和收缩的印尼航道开始25-17硕士(ref。gydF4y2Ba25gydF4y2Ba)和中美洲的航道(CAS)左右开始9.2 Ma (ref。gydF4y2Ba26gydF4y2Ba)。在模型试验中,中科院关闭已经观察到增加再生有限公司gydF4y2Ba2gydF4y2Ba存储在太平洋地区内部gydF4y2Ba27gydF4y2Ba,大概是增强suboxia在东太平洋。中科院的这个和其他影响关闭可能有助于解释在太平洋FB-δ增加gydF4y2Ba15gydF4y2BaN和ΔδgydF4y2Ba15gydF4y2BaNgydF4y2Ba便士gydF4y2Ba以及南大西洋FB-δ的相对稳定gydF4y2Ba15gydF4y2BaN后9.2 Ma(无花果。gydF4y2Ba2 b, cgydF4y2Ba)。然而,没有证据表明CAS的收缩到MMCO结束之后gydF4y2Ba26gydF4y2Ba。此外,即使CAS关闭开始之前,它不会解释太平洋FB-δ下降gydF4y2Ba15gydF4y2BaN和ΔδgydF4y2Ba15gydF4y2BaNgydF4y2Ba便士gydF4y2BaMMCO。最后,中科院关闭就不会EECO-associated变化有关。gydF4y2Ba
可能一些航道和/或盆地几何变化合谋产生观察MMCO和δEECO最小值gydF4y2Ba15gydF4y2BaNgydF4y2BaPgydF4y2Ba,δgydF4y2Ba15gydF4y2BaNgydF4y2Ba一个gydF4y2Ba和ΔδgydF4y2Ba15gydF4y2BaNgydF4y2Ba便士gydF4y2Ba。然而,δgydF4y2Ba15gydF4y2BaN的变化定义的最低在MMCO突然相对于构造变化,和他们配合ΔSSTgydF4y2BaL-HgydF4y2Ba(图的变化。gydF4y2Ba3gydF4y2Ba)。此外,MMCO内,有一个突然的δgydF4y2Ba15gydF4y2BaN增加现场872之际,恰逢米冰川作用在15.9 Ma(无花果。gydF4y2Ba3 cgydF4y2Ba),一个冷却事件没有任何已知的连接构造gydF4y2Ba28gydF4y2Ba。这些观察认为MMCO的脱氮下降和EECO居多的一个共同应对这些否则的温暖条件不同的时期。gydF4y2Ba

左边是MMCO和EECO在右边。gydF4y2Ba一个gydF4y2Ba,gydF4y2BabgydF4y2Ba编译,海温与底栖生物δgydF4y2Ba18gydF4y2BaO编译gydF4y2Ba8gydF4y2Ba,如无花果。gydF4y2Ba2gydF4y2Ba。这些数据表明,在气候最适条件,减少温度低纬度和高纬度地区地表水之间的区别。gydF4y2BacgydF4y2Ba,gydF4y2BadgydF4y2Ba,FB-δgydF4y2Ba15gydF4y2BaN核心网站(Δδ太平洋和大西洋之间的区别gydF4y2Ba15gydF4y2BaNgydF4y2Ba便士gydF4y2Ba),如在无花果。gydF4y2Ba2gydF4y2Ba。在gydF4y2BacgydF4y2Ba和gydF4y2BadgydF4y2Ba,其他的线表示δ的平均运行gydF4y2Ba18gydF4y2BaO底栖生物之间的区别和surface-dwelling planktic有孔虫(ΔδgydF4y2Ba18gydF4y2Ba在南大洋(O)gydF4y2BacgydF4y2BaODP网站1171年,红线gydF4y2Ba37gydF4y2Ba,gydF4y2Ba38gydF4y2Ba;gydF4y2BadgydF4y2Ba,ODP网站690年和738年,紫色线(ref。gydF4y2Ba41gydF4y2Ba和引用)。这些数据显示一个较弱的垂直密度梯度在南大洋气候最适条件,符合强通风南大洋深海的表面。gydF4y2BaegydF4y2Ba,gydF4y2BafgydF4y2Ba,FB-δgydF4y2Ba15gydF4y2BaN为这些时间间隔记录。在gydF4y2BaegydF4y2Ba的线表示平均记录导致FB-δgydF4y2Ba15gydF4y2BaN从ODP站点872(绿色)和DSDP网站516(蓝色)。孵化的酒吧在南大洋表明沉积间断,已被归因于更强的深海环流gydF4y2Ba39gydF4y2Ba。在gydF4y2BafgydF4y2Ba的线表示平均记录mixed-taxa FB-δgydF4y2Ba15gydF4y2BaN 1209年网站(浅绿色)和站点1263(浅蓝色)gydF4y2Ba18gydF4y2Ba,如无花果。gydF4y2Ba2gydF4y2Ba。新菌种FB-δgydF4y2Ba15gydF4y2BaN数据从网站516深蓝色。朋友,古新世;美国海洋,南大洋。gydF4y2Ba
全球海洋硝酸δgydF4y2Ba15gydF4y2BaN是强烈调制沉积比潜脱氮,收益率更高比率较低的δgydF4y2Ba15gydF4y2BaN (ref。gydF4y2Ba15gydF4y2Ba)。随着气候变暖,冰和其他进程的土地损失可能会导致海平面上升gydF4y2Ba12gydF4y2Ba,鼓励沉积脱氮gydF4y2Ba29日gydF4y2Ba。这种变化可能会认为低δ作出贡献gydF4y2Ba15gydF4y2BaN MMCO和EECO。然而,sea-level-driven沉积脱氮的变化会产生类似的δgydF4y2Ba15gydF4y2BaN下降在大西洋和太平洋盆地。因此,从极高的δ更大的下降gydF4y2Ba15gydF4y2BaN在872年和1209年网站不能完全解释增强沉积脱氮;相反,减少潜反硝化作用是必需的gydF4y2Ba18gydF4y2Ba。gydF4y2Ba
气候驱动ODZ收缩gydF4y2Ba
气候模型指出几个流程,努力缩小ODZs在全球变暖问题,提供可能的解释为我们找到减少潜MMCO和EECO脱氮。首先,减少upwelling-supported在赤道和热带太平洋生物出口生产将减少需氧量在地下gydF4y2Ba4gydF4y2Ba,gydF4y2Ba5gydF4y2Ba,gydF4y2Ba30.gydF4y2Ba。第二,海洋环流变化的室内,如增强南大洋深海形成,可能会增加氧气供应gydF4y2Ba3gydF4y2Ba,gydF4y2Ba6gydF4y2Ba。我们的特克斯gydF4y2Ba86年gydF4y2Ba基于重建的SST和ΔSSTgydF4y2BaL-HgydF4y2Ba机制(图是一致的。gydF4y2Ba2 a, bgydF4y2Ba)。一方面,全球气温和弱ΔSST就越高gydF4y2BaL-HgydF4y2Ba在MMCO和EECO应该削弱了热带大气环流gydF4y2Ba31日gydF4y2Ba,这将减少了赤道东太平洋上升流gydF4y2Ba4gydF4y2Ba。另一方面,加强深水形成等高纬度地区南大洋可能运送更多的热量从低到高纬度地区,进一步削弱ΔSSTgydF4y2BaL-HgydF4y2Ba(ref。gydF4y2Ba32gydF4y2Ba)。gydF4y2Ba
MMCO期间,赤道太平洋的重建水文学表明赤道太平洋温跃层的深化和压扁gydF4y2Ba33gydF4y2Ba应减少的营养供给引起的表面。这些发现符合减少低纬度生产和出口,因此,减少需氧量ODZ收缩的原因(无花果。gydF4y2Ba2 bgydF4y2Ba)。此外,模型研究gydF4y2Ba5gydF4y2Ba表明全球变暖的养分含量降低密度跃层水域通风热带地区,减少热带出口生产和导致ODZs合同。然而,证据改变热带出口生产整个MMCO EECO稀缺和模糊gydF4y2Ba34gydF4y2Ba。因此,尽管tropical-productivity-related解释是合理的,它仍然是未经证实的。gydF4y2Ba
了公海ODZs内密度跃层,跨度约150 - 1200米深度。因此,ODZs无疑是敏感的生物地球化学条件和循环在这些“中间”的深度gydF4y2Ba4gydF4y2Ba,gydF4y2Ba5gydF4y2Ba。然而,ODZs也受到潜在的深海,这水有助于他们通过垂直混合和上升流,特别是赤道附近和在海洋盆地的东部gydF4y2Ba30.gydF4y2Ba。在全球变暖的模拟预测海洋再氧化纪念和更长的时间尺度,这再氧化的结果更有活力的通风的深海,由增强对流在南大洋深处gydF4y2Ba3gydF4y2Ba,gydF4y2Ba6gydF4y2Ba。gydF4y2Ba
深海的氧浓度(以及整个海洋内部)与海洋紧密相连的“生物碳泵”,存储的呼吸系统有限公司gydF4y2Ba2gydF4y2Ba在深水域沉没,造成的后续补充矿质在表面水域产生的有机物质。更强的生物泵与更多的海洋有限公司gydF4y2Ba2gydF4y2Ba存储,降低大气COgydF4y2Ba2gydF4y2Ba深海,降低氧气浓度gydF4y2Ba35gydF4y2Ba。海洋生物泵的上限是由全球平均营养(磷酸或硝酸)浓度的海洋。然而,现代生物泵是低于这一限制,很大程度上是因为通风的南大洋深海gydF4y2Ba36gydF4y2Ba,这往往会填满海洋内部未使用的营养,减少耗氧量海洋内部由于“错失良机”nutrient-fuelled在南大洋表面生物生产力。因此,潜脱氮的下降在MMCO和EECO可能是由强大的南大洋深海的通风。gydF4y2Ba
这种可能性与其他的观察是一致的。MMCO期间,高分辨率的南大洋有孔虫氧同位素数据从ODP站点1171(参考文献。gydF4y2Ba37gydF4y2Ba,gydF4y2Ba38gydF4y2Ba)(古纬度约55°S)显示低planktic-benthic差异在MMCO和增加事件后,恰逢Δδ的增加gydF4y2Ba15gydF4y2BaNgydF4y2Ba便士gydF4y2Ba(无花果。gydF4y2Ba3 cgydF4y2Ba)。这些数据表明减少深海之间的密度梯度和南大洋表面,符合增加深度的颠覆和同时削弱全球生物泵。记录还显示更强的南大洋的一集密度分层(高ΔδgydF4y2Ba18gydF4y2BaOgydF4y2Babenthic-plankticgydF4y2Ba),与潜脱氮的增加(Δδ高gydF4y2Ba15gydF4y2BaNgydF4y2Ba便士gydF4y2Ba在米2)冰川作用约为15.9 Ma(无花果。gydF4y2Ba3 cgydF4y2Ba)。最后,在MMCO以及暂停在随后的冷却,最向极南大洋沉积物(ODP网站689年和690年)包含在沉积间断。这些中断与侵蚀事件引发的加强深水循环gydF4y2Ba39gydF4y2Ba,进一步支持加速深海通风是太平洋suboxia下降的一个原因(无花果。gydF4y2Ba3 c, egydF4y2Ba)。gydF4y2Ba
在EECO MMCO,氧同位素之间的梯度planktic和底栖有孔虫在高纬度地区的南大洋大幅减少gydF4y2Ba40gydF4y2Ba(无花果。gydF4y2Ba3 dgydF4y2Ba),又符合增加深推翻,恰逢ODZ收缩。此外,全球海洋重建垂直碳同位素的梯度表示在EECO疲软的全球生物泵gydF4y2Ba41gydF4y2Ba。这些观察结果支持的可能性增强南大洋推翻了两MMCO和EECO ODZ收缩。虽然我们一直关注南大洋推翻,北太平洋也表明了深水形成的新生代的温暖的时期gydF4y2Ba42gydF4y2Ba,gydF4y2Ba43gydF4y2Ba加强,可能有类似影响海洋生物泵和氧气。gydF4y2Ba
减少海洋生物碳泵的效率会增加大气COgydF4y2Ba2gydF4y2Ba在MMCO EECO。这种反馈,虽然假定缓冲上的“风化恒温器”过时间表gydF4y2Ba44gydF4y2Ba,可能会放大这些温暖的气候事件。Palaeobiogeochemical数据表明,现代南大洋深海的通风是减少在更新世冰河时代末期gydF4y2Ba45gydF4y2Ba,提供了一个解释低氧浓度的冰河时代深海相比间冰期和温暖的上新世gydF4y2Ba43gydF4y2Ba,gydF4y2Ba46gydF4y2Ba,gydF4y2Ba47gydF4y2Ba,gydF4y2Ba48gydF4y2Ba。这个解释更新世冰河周期意味着气候敏感性的南大洋深海的通风,符合我们的观察EECO MMCO,这表明,气候变暖会引起ocean-interior氧气浓度的增加。gydF4y2Ba
对未来气候gydF4y2Ba
我们的测量表明,在过去的时间的延长温暖,海洋suboxia是减少了,而不是扩大。这种变化的可能机制包括短期和长期的过程gydF4y2Ba3gydF4y2Ba,gydF4y2Ba4gydF4y2Ba,gydF4y2Ba5gydF4y2Ba,gydF4y2Ba6gydF4y2Ba,使得不确定性对未来几十年我们的研究结果是否有影响的全球变暖或长期。例如,如果一个热带太平洋生产力下降的主要原因是ODZ收缩在一个温暖的世界,然后收缩的缺氧区可能出现在未来几十年。相比之下,如果深海通风是占主导地位的原因,那么我们的研究结果只会成为重要的数百年来最早。无论如何,我们的结果表明,持续的趋势增强了公海suboxia报道gydF4y2Ba1gydF4y2Ba,gydF4y2Ba2gydF4y2Ba可能是数十年变化的结果吗gydF4y2Ba49gydF4y2Ba或瞬态响应全球变暖的速度,并不是一个永久的应对气候变暖。gydF4y2Ba
方法gydF4y2Ba
网站的核心位置gydF4y2Ba
新FB-δgydF4y2Ba15gydF4y2BaN进行分析是两个沉积物向极和阻碍利润的贫瘠副热带环流在大西洋(DSDP网站516)和太平洋(ODP网站872),分别。DSDP网站516(孔Z和F)(30°17′年代,35°17′W)钻1313米水深的深海钻探计划(DSDP)腿72在西南大西洋gydF4y2Ba50gydF4y2Ba。站点位于格兰德河上升。ODP网站872 (10°05′N, 162°51′E, 1095米水深)是空心的海山在赤道东太平洋的马绍尔群岛在144年大洋钻探计划(ODP)腿(ref。gydF4y2Ba51gydF4y2Ba)。疏松的沉积物包含丰富,保存完好的planktic有孔虫。我们的新数据编译之前FB-δ出版gydF4y2Ba15gydF4y2BaN太平洋ODP数据网站1209年和1263年大西洋ODP网站,其中大多数是复杂的类群gydF4y2Ba18gydF4y2Ba。此外,mixed-taxa FB-δgydF4y2Ba15gydF4y2BaN测量了几个样品中渐新世晚期和中新世早期年龄从太平洋ODP网站1211 (32°0′N, 157°51′E, 2907米水深),在靠近网站1209(扩展数据图。gydF4y2Ba1gydF4y2Ba)。过去的位置计算的基础上,参考。gydF4y2Ba52gydF4y2Ba,当现有估计在ODP报告gydF4y2Ba53gydF4y2Ba,gydF4y2Ba54gydF4y2Ba和图所示。gydF4y2Ba1gydF4y2Ba(地图使用海洋数据视图生成软件gydF4y2Ba55gydF4y2Ba和数据参考。gydF4y2Ba55gydF4y2Ba)。gydF4y2Ba
上面的表层水硝酸所有核心网站环境中完成消费,因此,应记录δgydF4y2Ba15gydF4y2BaN的硝酸盐提供密度跃层的表面。网站872年和1209年的边缘靠近nitrate-bearing地表水与赤道太平洋上升流有关。这可能修改FB-δgydF4y2Ba15gydF4y2Ba相对于δNgydF4y2Ba15gydF4y2BaN地下硝酸盐供应的地区,很可能增加FB-δgydF4y2Ba15gydF4y2BaN (ref。gydF4y2Ba56gydF4y2Ba)。因此,任何减少赤道上升流有时在过去可能FB-δ小幅减少gydF4y2Ba15gydF4y2BaN 872网站。gydF4y2Ba
我们也报告新的甘油二烷基甘油四醚(GDGTs)测量从DSDP网站516年和588年(26°07′年代,161°14′E, 1548米水深)以及ODP网站667 (4°34′N, 21°55′W, 3536米水深),704年(46 52°′年代,7°25′E, 2543米水深),730 (17°44′N, 57°42′E, 1071米水深),754(30°56′年代,93°34 E′1247米水深),1146 (19°27′N, 116°16′E, 2091米水深)和1263年(28°32′年代,2°46′E, 2717米水深)。新的测量允许我们生成一个连续的记录对海温的进化中低和高纬度地区新生代完全基于特克斯gydF4y2Ba86年gydF4y2BapalaeothermometergydF4y2Ba21gydF4y2Ba。gydF4y2Ba
年龄模型gydF4y2Ba
DSDP网站516中,我们使用一个组合年龄模型基于生物地层的论据gydF4y2Ba57gydF4y2Ba,gydF4y2Ba58gydF4y2Ba,gydF4y2Ba59gydF4y2Ba适应GTS12 (ref。gydF4y2Ba60gydF4y2Ba),磁性地层学Florindo et al。gydF4y2Ba61年gydF4y2Ba和年龄领带点基于K-Ar约会gydF4y2Ba62年gydF4y2Ba并为发生于k - pg边界铱飙升gydF4y2Ba63年gydF4y2Ba。ODP网站872年,我们使用了年龄模型基于生物地层的论据gydF4y2Ba64年gydF4y2Ba,gydF4y2Ba65年gydF4y2Ba适应GTS12时间表gydF4y2Ba60gydF4y2Ba。年龄模型基于生物地层学的ODP网站1209年和1263年在清晨里描述等。gydF4y2Ba18gydF4y2Ba。我们这些年龄模型修正天文调谐年表最近发表的Westerhold et al。gydF4y2Ba8gydF4y2Ba,gydF4y2Ba66年gydF4y2Ba。ODP网站1211的年龄模型是基于生物地层学gydF4y2Ba67年gydF4y2Ba。DSDP网站588的年龄模型是基于编译生物地层学、磁性地层学和同位素地层学(帕加尼et al。gydF4y2Ba68年gydF4y2Ba和引用)。ODP站点667岁从生物地层的绝对约束由线性插值(有孔虫和钙质超微化石)论据gydF4y2Ba69年gydF4y2Ba。ODP站点704岁被插值计算生物地层的(有孔虫和钙质超微化石)论据和磁性地层数据gydF4y2Ba70年gydF4y2Ba,通过调整新的特克斯gydF4y2Ba86年gydF4y2Ba记录转到特克斯gydF4y2Ba86年gydF4y2Ba记录从1171年网站和U1318中间中新世气候过渡,这是基于高分辨率的底栖生物δgydF4y2Ba18gydF4y2BaO地层学gydF4y2Ba71年gydF4y2Ba,gydF4y2Ba72年gydF4y2Ba。年龄对ODP网站730年由线性插值的生物地层的论据(有孔虫和钙质超微化石)gydF4y2Ba73年gydF4y2Ba。ODP网站754的年龄模型是基于生物地层学(钙质超微化石)gydF4y2Ba74年gydF4y2Ba。ODP网站1146的年龄模型是基于生物地层学(有孔虫)由内森和Leckie报道gydF4y2Ba75年gydF4y2Ba。年龄对ODP网站690/689是基于参考文献。gydF4y2Ba41gydF4y2Ba,gydF4y2Ba76年gydF4y2Ba。ODP站点1171岁是基于ref。gydF4y2Ba71年gydF4y2Ba。所有的生物地层的论据和古地磁的逆转被校准GTS12 (ref。gydF4y2Ba60gydF4y2Ba)(补充数据gydF4y2Ba3gydF4y2Ba和扩展数据图。gydF4y2Ba5gydF4y2Ba)。gydF4y2Ba
氮同位素分析gydF4y2Ba
大约600 - 800个人有孔虫的物种gydF4y2BaGlobigerinoides conglobatusgydF4y2Ba,gydF4y2BaTrilobatus sacculifergydF4y2Ba,gydF4y2BaOrbulina universagydF4y2Ba和gydF4y2BaSphaeroidinella dehiscensgydF4y2Ba选(250 - 400μm大小分数)中新世、上新世和更新世部分DSDP网站516和ODP网站872(扩展数据图。gydF4y2Ba1gydF4y2Ba)。gydF4y2BaDentoglobigerina altispiragydF4y2Ba和gydF4y2Bad . tripartitagydF4y2Ba均选用> 400 -μm大小分数。在516年的渐新世和始新世部分网站,我们测量genus-specific N同位素属gydF4y2BaDentoglobigerinagydF4y2Ba,gydF4y2BaTurborotaliagydF4y2Ba,gydF4y2BaAcarininagydF4y2Ba和gydF4y2BaSubbotinagydF4y2Ba以及混合有孔虫。的FB-δgydF4y2Ba15gydF4y2BaN在Martinez-Garcia实验室测量进行了马克斯·普朗克化学研究所(MPIC)。我们使用过硫酸盐FB-δoxidation-denitrifier方法gydF4y2Ba15gydF4y2BaN首次描述planktic有孔虫任et al。gydF4y2Ba17gydF4y2Ba,较小的调整协议。作为第一步,5 - 7毫克的有孔虫测试轻轻压碎,转移到一次性,15毫升聚丙烯,conical-base离心管和化学处理来消除外部污染。10毫升Na-polyphosphate解决方案(pH值8)添加到样品管,ultrasonicated 10年代去除粘土。与Milli-Q水冲洗样品三次后(通过填充和卸载),7毫升dithionite-citric酸溶液(Milli-Q 100毫升、柠檬酸钠6.2克、2 g碳酸氢钠,5 g连二亚硫酸钠,400μl 4 n氢氧化钠)被添加到每个样本gydF4y2Ba77年gydF4y2Ba。样品管被放置在80°C水浴30分钟。冷却后,所有样品都用Milli-Q水冲洗三次。接下来,执行一个氧化清洗步骤移除外部有机物:过硫酸钾/氢氧化钠溶液(2 g再结晶过硫酸钾,2 g氢氧化钠颗粒,100毫升Milli-Q水)添加和热压处理过的65分钟的125°C。冷却后,样本清洗四次Milli-Q水和干一夜之间在烤箱60°C。接下来,3 - 5毫克清洗外壳碎片重和盐酸溶解在45μl 4 n释放有机物进行分析。CaCO后gydF4y2Ba3gydF4y2Ba解散一步,有机N硝酸氧化是通过添加1毫升的第二个食谱的基本过硫酸钾溶液(0.7 g再结晶过硫酸钾,4毫升6.25 N氢氧化钠溶液,96毫升Milli-Q水)。硝酸浓度被转换为每个样品测量一氧化氮紧随其后的是化学发光检测gydF4y2Ba78年gydF4y2Ba。随后,5 nmol N样品溶液的硝酸使用脱氮剂方法,转化为一氧化二氮和δgydF4y2Ba15gydF4y2BaN被气体chromatography-isotope比率质谱测量gydF4y2Ba79年gydF4y2Ba,gydF4y2Ba80年gydF4y2Ba,gydF4y2Ba81年gydF4y2Ba,gydF4y2Ba82年gydF4y2Ba。与先前的研究gydF4y2Ba19gydF4y2Ba,gydF4y2Ba29日gydF4y2Ba,gydF4y2Ba83年gydF4y2Ba,gydF4y2Ba84年gydF4y2Ba之前,样品没有pH值调整注入反硝化细菌;相反,他们没有调整注入2.75毫升媒体缓冲pH值6.3和包含反硝化细菌(见参考。gydF4y2Ba82年gydF4y2Ba,gydF4y2Ba85年gydF4y2Ba详情)。量化修正同位素值的精度和准确性,为每个系列的30个样品,总共九个不同的内部(MPIC)有孔虫和珊瑚实验室标准进行分析。以下分析(各一式三份):珊瑚标准分类单元gydF4y2BaPoritesgydF4y2Ba和δ(PO-1)gydF4y2Ba15gydF4y2BaN为6.2±0.3‰,珊瑚分类单元的标准gydF4y2BaLopheliagydF4y2Ba和δgydF4y2Ba15gydF4y2Ba10.1±0.4‰(ref的N。gydF4y2Ba85年gydF4y2Ba)和混合有孔虫标准(63 - 315μm大小分数)从北大西洋(MSM58-17-1)δgydF4y2Ba15gydF4y2BaN的5.84‰(没有评估的不确定性)。校准后硝酸与国际同位素参考IAEA-NO3 USGS-34和氧化空白校正,有孔虫的长期分析精度标准为0.23‰(1个标准差,gydF4y2BangydF4y2Ba= 22)。每个氧化氧化空白样本通常低于0.2 nmol N。gydF4y2Ba
分析GDGTsgydF4y2Ba
5 - 20 g冻干沉积物样品的提取和分离成两个分数MPIC Martinez-Garcia实验室,之后提出的协议Auderset et al。gydF4y2Ba86年gydF4y2Ba。加速溶剂萃取后冷冻沉积物样品,我们添加了60μl内部标准(CgydF4y2Ba46gydF4y2Ba-GDGT,合成了Patwardhan和汤普森gydF4y2Ba87年gydF4y2Ba为量化)。提取干在离心火箭蒸发器(Genevac)和聚四氟乙烯过滤器过滤(0.2 -μm孔隙大小)己烷的混合物:1.4%异丙醇(十六进制:异丙醇)。GDGTs分析使用高效液相色析法(安捷伦1260∞)耦合single-quadrupole质谱仪探测器(安捷伦,6130)提出的协议Hopmans et al。gydF4y2Ba88年gydF4y2Ba,有些小的修改。正常阶段分离实现了一个超高性能的液体色析法硅胶柱(本·HILIC列,2.1毫米×150毫米,1.7μm;水域)保持在30°C。流量的1.4%十六进制:音标流动相为0.2毫升mingydF4y2Ba−1gydF4y2Ba和保持不变的前25分钟,紧随其后的是一个梯度十六进制:3.5%异丙醇在25分钟和10%异丙醇在己烷column-cleaning一步。我们使用一个单离子监测质量gydF4y2Ba米gydF4y2Ba/gydF4y2BazgydF4y2Ba= 1302。3,1300。3、1298 .3 1296。3,1292。3,744.0,1050 .0,1036 .0,1022 0,1020 0和1018 0。注射量是5μl或20μl,根据样品的浓度分析。gydF4y2Ba
太平洋和大西洋FB-δgydF4y2Ba15gydF4y2BaN栈gydF4y2Ba
由于的出现和灭绝planktic有孔虫类群随着时间的推移,重要的是要评估FB-δ之间的关系gydF4y2Ba15gydF4y2BaN的灭绝有孔虫的现代分类单元的合并N同位素信号进行了研究gydF4y2Ba19gydF4y2Ba,gydF4y2Ba89年gydF4y2Ba,gydF4y2Ba90年gydF4y2Ba。我们的新数据集从DSDP站点516表明FB-δgydF4y2Ba15gydF4y2BaN的gydF4y2Bat . sacculifergydF4y2Ba、现代photosymbiotic surface-dwelling物种,与FB-δ重叠gydF4y2Ba15gydF4y2BaN已经灭绝的物种gydF4y2Ba美国dehiscensgydF4y2Ba,以及FB-δgydF4y2Ba15gydF4y2BaN混合planktic有孔虫在中新世和晚渐新世早期(扩展数据图。gydF4y2Ba1gydF4y2Ba)。此外,在渐新世和始新世时期,FB-δgydF4y2Ba15gydF4y2BaN的属gydF4y2BaDentoglobigerinagydF4y2Ba,gydF4y2BaTurborotaliagydF4y2Ba和gydF4y2BaAcarininagydF4y2Ba重叠的gydF4y2Ba美国dehiscensgydF4y2Ba和混合物种,这表明没有明显的δgydF4y2Ba15gydF4y2Ba现代和已经灭绝的物种有孔虫N之间的偏移量。在872年ODP的网站,gydF4y2Bao . universagydF4y2Ba和gydF4y2Bat . sacculifergydF4y2Ba,两个现代photosymbiotic表面居民,显示非常相似的值在过去15最高产量研究。此外,我们的数据显示,facebookgydF4y2Ba- - - - - -gydF4y2BaδgydF4y2Ba15gydF4y2Ba这两个现代物种的N和共病的重叠gydF4y2Bad . altispiragydF4y2Ba通过中新世和上新世时期,尽管大量(但一致)N含量的差异。gydF4y2BaDentoglobigerina altispiragydF4y2BaδgydF4y2Ba15gydF4y2Ba反过来,N值是相似的gydF4y2Bad . tripartitagydF4y2Ba在中新世早期。通过这种方式,我们可以从不同的物种有孔虫组合测量/属获得连续FB-δ的记录gydF4y2Ba15gydF4y2BaN在整个新生代(扩展数据图。gydF4y2Ba1gydF4y2Ba)。的FB-δgydF4y2Ba15gydF4y2BaN大西洋和太平洋栈在无花果。gydF4y2Ba2摄氏度gydF4y2Ba平均的菌种,genus-specific和mixed-taxa值扩展数据图所示。gydF4y2Ba1gydF4y2Ba。正如下面所讨论的,这股ODP网站1209迁移以进入北太平洋副热带环流在40最高产量研究(无花果。gydF4y2Ba1gydF4y2Ba),所以ODP网站1209记录被截断在这个年龄。gydF4y2Ba
核心网站迁移和FB-δgydF4y2Ba15gydF4y2BaNgydF4y2Ba
海底迁移是一个无处不在的担忧Cenozoic-long palaeoceanographic重建。网站1209年认股权证在这方面考虑,因为它从ODZ-influenced热带北太平洋中部迁移到西部北太平洋副热带环流之间的早期和mid-Cenozoic(无花果。gydF4y2Ba1gydF4y2Ba)。几个观测表明FB-δ大幅下滑gydF4y2Ba15gydF4y2BaN在EECO不是由于网站迁移。首先,FB-δgydF4y2Ba15gydF4y2BaN下降到EECO也发生在北和南大西洋,在网站迁移不会有这样的效果gydF4y2Ba18gydF4y2Ba(无花果。gydF4y2Ba1gydF4y2Ba)。第二,FB-δgydF4y2Ba15gydF4y2BaN类似规模的下降是EECO MMCO,认为相同的动态工作对温暖的事件。第三,FB-δgydF4y2Ba15gydF4y2BaN 1209网站显示的变化是相反的网站迁移的意义:一个大约6‰δgydF4y2Ba15gydF4y2BaN增加从晚白垩世古新世和一个大约3‰增加约40 Ma (ref。gydF4y2Ba18gydF4y2Ba)。然而,网站1209的敏感性脱氮的同位素信号在东部热带太平洋最终被迁移,减少之间的变化发生中始新世和早中新世。这是由早期中新世FB-δ的相似性表示gydF4y2Ba15gydF4y2BaN 1211网站(网站1209)的亲密邻居FB-δgydF4y2Ba15gydF4y2Ba516年在南大西洋,这两个比FB-δ大幅降低gydF4y2Ba15gydF4y2BaN在太平洋网站872在中新世早期(扩展数据图。gydF4y2Ba1gydF4y2Ba)。这促使我们使用网站872年从渐新世。gydF4y2Ba
纬向风场栈gydF4y2Ba
呈现纬向风场的叠加,我们总共有653新特克斯gydF4y2Ba86年gydF4y2Ba风场测量低(ODP网站667年和730年)中间(DSDP网站516年和588年,ODP网站754年,1146年和1263年)和高纬度地区(ODP站点704)与现有的特克斯和集成的记录gydF4y2Ba86年gydF4y2Ba风场数据集(包括4474数据点53网站和48出版物)覆盖过去69 Ma(参考文献。gydF4y2Ba18gydF4y2Ba,gydF4y2Ba72年gydF4y2Ba,gydF4y2Ba91年gydF4y2Ba,gydF4y2Ba92年gydF4y2Ba,gydF4y2Ba93年gydF4y2Ba,gydF4y2Ba94年gydF4y2Ba,gydF4y2Ba95年gydF4y2Ba,gydF4y2Ba96年gydF4y2Ba,gydF4y2Ba97年gydF4y2Ba,gydF4y2Ba98年gydF4y2Ba,gydF4y2Ba99年gydF4y2Ba,gydF4y2BaOne hundred.gydF4y2Ba,gydF4y2Ba101年gydF4y2Ba,gydF4y2Ba102年gydF4y2Ba,gydF4y2Ba103年gydF4y2Ba,gydF4y2Ba104年gydF4y2Ba,gydF4y2Ba105年gydF4y2Ba,gydF4y2Ba106年gydF4y2Ba,gydF4y2Ba107年gydF4y2Ba,gydF4y2Ba108年gydF4y2Ba,gydF4y2Ba109年gydF4y2Ba,gydF4y2Ba110年gydF4y2Ba,gydF4y2Ba111年gydF4y2Ba,gydF4y2Ba112年gydF4y2Ba,gydF4y2Ba113年gydF4y2Ba,gydF4y2Ba114年gydF4y2Ba,gydF4y2Ba115年gydF4y2Ba,gydF4y2Ba116年gydF4y2Ba,gydF4y2Ba117年gydF4y2Ba,gydF4y2Ba118年gydF4y2Ba,gydF4y2Ba119年gydF4y2Ba,gydF4y2Ba120年gydF4y2Ba,gydF4y2Ba121年gydF4y2Ba,gydF4y2Ba122年gydF4y2Ba,gydF4y2Ba123年gydF4y2Ba,gydF4y2Ba124年gydF4y2Ba,gydF4y2Ba125年gydF4y2Ba,gydF4y2Ba126年gydF4y2Ba,gydF4y2Ba127年gydF4y2Ba,gydF4y2Ba128年gydF4y2Ba,gydF4y2Ba129年gydF4y2Ba,gydF4y2Ba130年gydF4y2Ba,gydF4y2Ba131年gydF4y2Ba,gydF4y2Ba132年gydF4y2Ba,gydF4y2Ba133年gydF4y2Ba,gydF4y2Ba134年gydF4y2Ba,gydF4y2Ba135年gydF4y2Ba,gydF4y2Ba136年gydF4y2Ba)。总结了数据和引用补充数据gydF4y2Ba2gydF4y2Ba和扩展数据无花果所示。gydF4y2Ba2gydF4y2Ba和gydF4y2Ba3gydF4y2Ba。我们分类根据年龄调整古纬度的不同数据集使用paleolatitude.orggydF4y2Ba52gydF4y2Ba。分类如下:高纬度地区(高于50°N / S),低纬度(0-20°N / S),中纬度(20 - 40°N / S),过渡(40 - 50°N / S)和肋上升流区域(扩展数据图。gydF4y2Ba3gydF4y2Ba)。不幸的是,低纬度网站稀缺和海温记录不提供一个连续的新生代。中纬度记录海温趋势和绝对值更加丰富和展示非常类似于低纬度的间隔都记录重叠。因此,我们结合中间/记录计算低纬度地区海温纬向风场梯度。在无花果。gydF4y2Ba2gydF4y2Ba,我们估计之间的梯度和中/低高纬度(ΔSSTgydF4y2BaL-HgydF4y2Ba= SSTgydF4y2Ba中/低gydF4y2Ba−SSTgydF4y2Ba高gydF4y2Ba)在应用本地回归(黄土)平滑系数为0.02和0.2最高产量研究抽样步骤(使用Python代码可以在GitHub:gydF4y2Bahttps://github.com/audersea/Auderset_etal_2022_naturegydF4y2Ba)。沉积物岩心位于过渡区域(40°之间的古纬度和50°N / S),以及upwelling-sensitive位置(ODP 1085, ODP 1087和ODP 846),没有考虑这些计算本地更改的上升流的影响减到最小强度和额迁移。gydF4y2Ba
估计SST特克斯gydF4y2Ba86年gydF4y2Ba金提出的价值观,我们使用校准等。gydF4y2Ba137年gydF4y2Ba(特克斯gydF4y2BaHgydF4y2Ba86年gydF4y2Ba)和蒂尔尼TingleygydF4y2Ba138年gydF4y2Ba(特克斯gydF4y2Ba86年BayspargydF4y2Ba,但有以下参数:prior_mean = 28;prior_std = 10;托尔= 0.15;n_samp = 2500)。总的来说,获得的趋势在计算校准的纬向温度梯度是相同的,但是有稍大的振幅的变化Bayspar-derived梯度(扩展数据图。gydF4y2Ba4 bgydF4y2Ba)。此外,我们也估计纬向梯度使用原始特克斯gydF4y2Ba86年gydF4y2Ba我们编译和将它比作δ值gydF4y2Ba15gydF4y2BaN梯度(扩展数据图。gydF4y2Ba4摄氏度gydF4y2Ba)。好原始特克斯之间的相关性gydF4y2Ba86年gydF4y2Ba梯度和δgydF4y2Ba15gydF4y2BaN梯度表明这是一个健壮的功能独立于所使用的校准。最后,我们比较重建的纬向风场梯度Cramwinckel等提出的一个。gydF4y2Ba99年gydF4y2Ba58和之间的时间间隔30 Ma,基于底栖生物的结合氧同位素记录高纬度和特克斯gydF4y2Ba86年gydF4y2Ba低纬度地区重建(扩展数据图。gydF4y2Ba4摄氏度gydF4y2Ba)。重构吻合较好;然而,这种方法Cramwinckel等紧随其后。gydF4y2Ba99年gydF4y2Ba只适用于没有实质性贡献的时期大陆冰底栖生物δgydF4y2Ba18gydF4y2BaO信号,限制其应用后Eocene-Oligocene边界。gydF4y2Ba
提出了几个指标来评估潜在来源的变化类异戊二烯GDGTsgydF4y2Ba110年gydF4y2Ba,gydF4y2Ba139年gydF4y2Ba,gydF4y2Ba140年gydF4y2Ba,gydF4y2Ba141年gydF4y2Ba。我们报告中对这些指标的值补充数据gydF4y2Ba2gydF4y2Ba。根据先前的研究表明,我们排除GDGTs数据具有高甲烷指数(0.4以上)和高GDGTgydF4y2BaRSgydF4y2Ba(30岁以上)从我们的计算纬度的温度梯度(355个样本)(扩展数据图。gydF4y2Ba3gydF4y2Ba)。分支和类异戊二烯四醚(位)指数提出了估计的相对贡献soil-derived GDGTs水生沉积环境特点是大型输入从附近的土壤(即肋海洋沉积物和湖泊沉积物)gydF4y2Ba142年gydF4y2Ba。在这些环境中,已经提出一些值低于0.4表明陆地GDGTs小投入,因此,一个微不足道的影响陆地GDGTs估计海洋上层温度gydF4y2Ba139年gydF4y2Ba。位的值在0.4以上样品通常不用于古温度计算由于潜在的偏见,土壤GDGTs输入可能对估计SST介绍。然而,这个索引来估算陆地输入的适用性GDGTs公海设置受到质疑,因为(1)支GDGTs可以产生了原位gydF4y2Ba143年gydF4y2Ba,gydF4y2Ba144年gydF4y2Ba,gydF4y2Ba145年gydF4y2Ba,gydF4y2Ba146年gydF4y2Ba,gydF4y2Ba147年gydF4y2Ba和(2)退化率crenarchaeol是支GDGTs的两倍gydF4y2Ba148年gydF4y2Ba。因此,在某些环境中,可以高一些指数(0.4以上),尽管brGDGTs陆地输入相对较低。在这些情况下,样本与> 0.4已经使用了古温度重建gydF4y2Ba71年gydF4y2Ba,gydF4y2Ba117年gydF4y2Ba,gydF4y2Ba129年gydF4y2Ba。近年来,它已被证明在situ-produced brGDGTs往往表现为循环brGDGTs的丰度较高,导致高值#戒指gydF4y2Ba脂鲤gydF4y2Ba指数,有人建议#戒指gydF4y2Ba脂鲤gydF4y2Ba值> 0.7表明主要原位支GDGTs的生产gydF4y2Ba149年gydF4y2Ba。一些新的样品报告,以及一些样品发表在先前的研究中包含我们的编译,有点值> 0.4。然而,一些样品的位的值> 0.4 #戒指也值gydF4y2Ba脂鲤gydF4y2Ba指数> 0.7,这表明brGDGTs原位产生的,而不是从陆地运输的土壤。在纬向风场梯度的计算,我们排除样品位> 0.4如果#戒指gydF4y2Ba脂鲤gydF4y2Ba< 0.7(178个样本),以避免潜在的影响陆地GDGTs估计SST(见扩展数据图。gydF4y2Ba3gydF4y2Ba补充数据gydF4y2Ba2gydF4y2Ba和代码gydF4y2Bahttps://github.com/audersea/paper_2022_naturegydF4y2Ba)。然而,我们注意到,没有实质性的差异对海温梯度如果所有的样本与钻头> 0.4中包括计算(补充数据gydF4y2Ba2gydF4y2Ba),这表明陆地上GDGTs估计SST的潜在影响可能是最少的,尽管高值。gydF4y2Ba
数据可用性gydF4y2Ba
数据集在补充数据是可用的gydF4y2Ba1gydF4y2Ba- - - - - -gydF4y2Ba3gydF4y2Ba和泛大陆数据库(gydF4y2Bahttps://doi.org/10.1594/PANGAEA.943130gydF4y2Ba)。gydF4y2Ba
代码的可用性gydF4y2Ba
代码用于滤波、平滑和梯度计算对海温和δgydF4y2Ba15gydF4y2BaN数据集可以在GitHub (gydF4y2Bahttps://github.com/audersea/Auderset_etal_2022_naturegydF4y2Ba)。gydF4y2Ba
引用gydF4y2Ba
Stramma, L。,Johnson, G. C., Sprintall, J. & Mohrholz, V. Expanding oxygen-minimum zones in the tropical oceans.科学gydF4y2Ba320年gydF4y2Ba,655 - 658 (2008)。gydF4y2Ba
Schmidtko, S。,Stramma, L。&Visbeck, M. Decline in global oceanic oxygen content during the past five decades.自然gydF4y2Ba542年gydF4y2Ba,335 - 339 (2017)。gydF4y2Ba
山本,a . et al。全球深海氧化增强通风在南大洋长期全球变暖。gydF4y2Ba水珠。Biogeochem。周期gydF4y2Ba29日gydF4y2Ba,1801 - 1815 (2015)。gydF4y2Ba
Takano Y。伊藤,T。&多伊奇,C。Projected centennial oxygen trends and their attribution to distinct ocean climate forcings.水珠。Biogeochem。周期gydF4y2Ba32gydF4y2Ba,1329 - 1349 (2018)。gydF4y2Ba
傅,W。,Primeau, F., Keith Moore, J., Lindsay, K. & Randerson, J. T. Reversal of increasing tropical ocean hypoxia trends with sustained climate warming.水珠。Biogeochem。周期gydF4y2Ba32gydF4y2Ba,551 - 564 (2018)。gydF4y2Ba
Frolicher, t . et al .对比上和深海氧反应旷日持久的全球变暖。gydF4y2Ba水珠。Biogeochem。周期gydF4y2Ba34gydF4y2Bae2020GB006601 (2020)。gydF4y2Ba
基林,r F。,Körtzinger, A. & Gruber, N. Ocean deoxygenation in a warming world.为基础。启3月科学。gydF4y2Ba2gydF4y2Ba,199 - 229 (2010)。gydF4y2Ba
Westerhold, t . et al。一个天文数字般的约会记录地球的气候和其可预测性在过去的6600万年。gydF4y2Ba科学gydF4y2Ba369年gydF4y2Ba,1383 - 1387 (2020)。gydF4y2Ba
汉森,J。佐藤,M。,Russell, G. & Kharecha, P. Climate sensitivity, sea level and atmospheric carbon dioxide.费罗斯。反式。r . Soc。一个数学。理论物理。Eng。科学。gydF4y2Ba371年gydF4y2Ba,20120294 (2013)。gydF4y2Ba
Anagnostou、大肠等。改变大气COgydF4y2Ba2gydF4y2Ba新生代早期气候的浓度是主要推动力。gydF4y2Ba自然gydF4y2Ba533年gydF4y2Ba,380 - 384 (2016)。gydF4y2Ba
Sosdian, s m . et al .制约新第三纪的进化海洋碳酸盐岩化学使用硼同位素的pH值代理。gydF4y2Ba地球的星球。科学。列托人。gydF4y2Ba498年gydF4y2Ba,362 - 376 (2018)。gydF4y2Ba
米勒,k . g . et al .新生代海平面和冰冻圈从深海地球化学演化和大陆边缘记录。gydF4y2Ba科学。睡觉。gydF4y2Ba6gydF4y2Baeaaz1346 (2020)。gydF4y2Ba
伯克,k·d . et al .上新世和始新世为不远的未来气候条件下提供最好的类似物。gydF4y2BaProc。《科学。gydF4y2Ba115年gydF4y2Ba,13288 - 13293 (2018)。gydF4y2Ba
Cline, j . &卡普兰,即硝酸溶解在脱氮同位素分馏的北太平洋东部的热带。gydF4y2Ba3月化学。gydF4y2Ba3gydF4y2Ba,271 - 299 (1975)。gydF4y2Ba
迪沃Brandes j . A & A . h .一个全球海洋固定氮同位素预算:对全新世氮循环的影响。gydF4y2Ba水珠。Biogeochem。周期gydF4y2Ba16gydF4y2Ba,67-1-67-14 (2002)。gydF4y2Ba
Altabet, m . a . et al .沉没的氮同位素生物地球化学粒子从北太平洋东部的边缘。gydF4y2Ba深海研究》第一部分Oceanogr。人民行动党>,gydF4y2Ba46gydF4y2Ba,655 - 679 (1999)。gydF4y2Ba
任,h . et al。有孔虫减少氮的同位素证据固定在冰河时代大西洋。gydF4y2Ba科学gydF4y2Ba323年gydF4y2Ba,244 - 248 (2009)。gydF4y2Ba
清晨,e . r . et al .氮同位素证据扩大海洋suboxia新生代早期。gydF4y2Ba科学gydF4y2Ba364年gydF4y2Ba,386 - 389 (2019)。gydF4y2Ba
任,H。,Sigman, D. M., Thunell, R. C. & Prokopenko, M. G. Nitrogen isotopic composition of planktonic foraminifera from the modern ocean and recent sediments.Limnol。Oceanogr。gydF4y2Ba57gydF4y2Ba,1011 - 1024 (2012)。gydF4y2Ba
西格曼,d m & Fripiat fgydF4y2Ba海洋科学百科全书gydF4y2Ba第三版(eds柯克科克伦,J。,Bokuniewicz, H. J. & Yager, P. L.) 263–278 (Academic, 2019).
思,S。,Hopmans, e . C。,Schefuß, E. & Damste, J. S. S. Distributional variations in marine crenarchaeotal membrane lipids: a new tool for reconstructing ancient sea water temperatures?地球的星球。科学。列托人。gydF4y2Ba204年gydF4y2Ba,265 - 274 (2002)。gydF4y2Ba
Bindoff: l .等人gydF4y2Ba政府间气候变化专门委员会的特别报道,海洋和冰冻圈变化的气候gydF4y2Ba(eds Portner H.-O。et al . Ch。5, 477 - 587(剑桥大学出版社,2022)。gydF4y2Ba
上演,o . M。弗兰克,M。,Betzler, C., Zammit, R. & Waldmann, N. D. Two-step closure of the Miocene Indian Ocean Gateway to the Mediterranean.科学。代表。gydF4y2Ba9gydF4y2Ba8842 (2019)。gydF4y2Ba
Lagabrielle Y。Godderis, Y。,Donnadieu, Y., Malavieille, J. & Suarez, M. The tectonic history of Drake Passage and its possible impacts on global climate.地球的星球。科学。列托人。gydF4y2Ba279年gydF4y2Ba,197 - 211 (2009)。gydF4y2Ba
Kuhnt, W。,Holbourn, A., Hall, R., Zuvela, M. & Käse, R. in在东亚陆缘海Continent-Ocean交互gydF4y2Ba卷149 (eds绝壁,P。Kuhnt, W。,Wang, P. & Hayes, D.) 299–320 (American Geophysical Union, 2004).
O 'Dea a . et al .巴拿马地峡的形成。gydF4y2Ba科学。睡觉。gydF4y2Ba2gydF4y2Bae1600883 (2016)。gydF4y2Ba
长袋网,j·G。,D’Orgeville, M. & Weaver, A. J. Drake Passage and Central American Seaway controls on the distribution of the oceanic carbon reservoir.水珠。星球。改变gydF4y2Ba128年gydF4y2Ba,72 - 82 (2015)。gydF4y2Ba
米勒,k . G。,Wright, J. D. & Fairbanks, R. G. Unlocking the ice house: Oligocene‐Miocene oxygen isotopes, eustasy, and margin erosion.j .地球物理学。研究固体地球gydF4y2Ba96年gydF4y2Ba,6829 - 6848 (1991)。gydF4y2Ba
任,h . et al .冰期/间冰期影响海平面变化对海洋氮循环。gydF4y2BaProc。《科学。gydF4y2Ba114年gydF4y2BaE6759-E6766 (2017)。gydF4y2Ba
格纳纳德西肯。,Russell, J. & Zeng, F. How does ocean ventilation change under global warming.海洋科学。gydF4y2Ba3gydF4y2Ba,利润率达到(2007)。gydF4y2Ba
Vecchi, g . a . et al .减弱的热带太平洋由于人为强迫大气环流。gydF4y2Ba自然gydF4y2Ba441年gydF4y2Ba,73 - 76 (2006)。gydF4y2Ba
道格拉斯,p . m . et al .明显的纬向异质性在始新世南部高纬度海洋表面温度。gydF4y2BaProc。《科学。美国gydF4y2Ba111年gydF4y2Ba,6582 - 6587 (2014)。gydF4y2Ba
松井,h . et al .垂直热梯度历史在赤道东太平洋早中新世中期:影响赤道温跃层的发展。gydF4y2Ba古海洋学gydF4y2Ba32gydF4y2Ba,729 - 743 (2017)。gydF4y2Ba
万登伯格,m . d . & Jarrard r . d .新生代大规模积累利率在赤道太平洋基于高分辨率海洋钻探计划的矿物学腿199。gydF4y2Ba古海洋学gydF4y2Ba19gydF4y2BaPA2021 (2004)。gydF4y2Ba
布勒克,w·s·冰川时期海洋化学。gydF4y2BaGeochim。Cosmochim。学报gydF4y2Ba46gydF4y2Ba,1689 - 1705 (1982)。gydF4y2Ba
Sarmiento, j·l . & Toggweiler j .的新模式决定大气pCO海洋的作用gydF4y2Ba2gydF4y2Ba。gydF4y2Ba自然gydF4y2Ba308年gydF4y2Ba,621 - 624 (1984)。gydF4y2Ba
Shevenell a E。,Kennett, J. P. & Lea, D. W. Middle Miocene southern ocean cooling and Antarctic cryosphere expansion.科学gydF4y2Ba305年gydF4y2Ba,1766 - 1770 (2004)。gydF4y2Ba
Shevenell a E。,Kennett, J. P. & Lea, D. W. Southern Ocean Middle Miocene ODP1171 Foraminifer Stable Isotope and Mg/Ca Data. IGBP PAGES/World Data Center for Paleoclimatology, Data Contribution Series # 2006-061, NOAA/NCDC Paleoclimatology Program (2006).
赖特,k . g . j . d . &米勒gydF4y2Ba南极的古环境:全球变化的角度来看,第二部分gydF4y2Ba(eds, j . p . & Warnke d . a) 1 - 25(美国地球物理学联合会,1993)。gydF4y2Ba
施耐德,l . J。,Bralower, t·J。&Kump, L. R. Response of nannoplankton to early Eocene ocean destratification.Palaeogeogr。Palaeoclimatol。Palaeoecol。gydF4y2Ba310年gydF4y2Ba,152 - 162 (2011)。gydF4y2Ba
柄,a K。,Kump, L. R. & Bralower, T. J. Variations in the oceanic vertical carbon isotope gradient and their implications for the Paleocene‐Eocene biological pump.古海洋学gydF4y2Ba23gydF4y2BaPA3222 (2008)。gydF4y2Ba
黑格,a . m . et al .对流在新生代早期北太平洋的深水。gydF4y2Ba地质gydF4y2Ba40gydF4y2Ba,527 - 530 (2012)。gydF4y2Ba
节:j . et al .活跃太平洋经向翻转环流(PMOC)在温暖的上新世。gydF4y2Ba科学。睡觉。gydF4y2Ba3gydF4y2Bae1700156 (2017)。gydF4y2Ba
沃克,j . C。,H一个ys, P. & Kasting, J. F. A negative feedback mechanism for the long‐term stabilization of Earth’s surface temperature.j .地球物理学。海洋》gydF4y2Ba86年gydF4y2Ba,9776 - 9782 (1981)。gydF4y2Ba
西格曼,d . m . et al .南大洋在冰河时代:回顾南极表面隔离假说,北太平洋的比较。gydF4y2Ba皮疹。科学。牧师。gydF4y2Ba254年gydF4y2Ba106732 (2021)。gydF4y2Ba
弗朗索瓦,r . et al。南大洋的贡献马莲分层低大气COgydF4y2Ba2gydF4y2Ba浓度在上次冰河时代。gydF4y2Ba自然gydF4y2Ba389年gydF4y2Ba,929 - 935 (1997)。gydF4y2Ba
Jaccard, s . L。,Galbraith, E. D., Martínez-García, A. & Anderson, R. F. Covariation of deep Southern Ocean oxygenation and atmospheric CO2gydF4y2Ba在最后一个冰河时代。gydF4y2Ba自然gydF4y2Ba530年gydF4y2Ba,207 - 210 (2016)。gydF4y2Ba
Hoogakker, b . a . et al .冰川扩张热带东太平洋的海水缺氧。gydF4y2Ba自然gydF4y2Ba562年gydF4y2Ba,410 - 413 (2018)。gydF4y2Ba
多伊奇,C。白利糖度,H。伊藤,T。,Frenzel, H. & Thompson, L. Climate-forced variability of ocean hypoxia.科学gydF4y2Ba333年gydF4y2Ba,336 - 339 (2011)。gydF4y2Ba
巴克,p . f . et al . 516年网站:格兰德河上升。深海钻探计划的初步报告,72卷,155 - 338。gydF4y2Bahttps://doi.org/10.2973/dsdp.proc.72.105.1983gydF4y2Ba(美国政府印刷局,1980)。gydF4y2Ba
我Premoli席尔瓦。,H一个ggerty, J. & Rack, F. Site 872. Proceedings of the Ocean Drilling Program, Initial Reports, Vol. 144, 105–144.https://doi.org/10.2973/odp.proc.ir.143.105.1993gydF4y2Ba(德州农工大学,1993)。gydF4y2Ba
van Hinsbergen, d . j . et al .古气候研究paleolatitude计算器。gydF4y2Ba《公共科学图书馆•综合》gydF4y2Ba10gydF4y2Bae0126946 (2015)。gydF4y2Ba
Bralower, t·J。,我Premoli席尔瓦。&Malone, M. Leg 198 summary. Proceedings of the Ocean Drilling Program, Initial Reports, Vol. 198, 1–148.https://doi.org/10.2973/odp.proc.ir.198.101.2002gydF4y2Ba(德州农工大学,2002)。gydF4y2Ba
啤酒,r与海洋数据视图的数据分析和可视化。gydF4y2BaCMOS公牛。SCMOgydF4y2Ba43gydF4y2Ba第四,(2015)。gydF4y2Ba
加西亚,h . et al . 2018年世界海洋图集:产品文档。a . Mishonov技术编辑(2019)。gydF4y2Ba
科斯塔k . m . et al .没有铁施肥赤道太平洋在最后一个冰河时代。gydF4y2Ba自然gydF4y2Ba529年gydF4y2Ba,519 - 522 (2016)。gydF4y2Ba
伯格伦,w。,一个ubry, M. P. & Hamilton, N. in Neogene magnetobiostratigraphy of Deep Sea Drilling Project Site 516 (Rio Grande Rise, South Atlantic). Initial Reports of the Deep Sea Drilling Project, Vol. 72, 675–706.https://doi.org/10.2973/dsdp.proc.72.130.1983gydF4y2Ba(美国政府印刷局,1983)。gydF4y2Ba
赫斯,J。,Stott, L. D., Bender, M. L., Kennett, J. P. & Schilling, J. G. The Oligocene marine microfossil record: age assessments using strontium isotopes.古海洋学gydF4y2Ba4gydF4y2Ba,655 - 679 (1989)。gydF4y2Ba
魏、w &小智慧,s . w .早第三纪钙质超微化石magnetobiochronology:从南大西洋DSDP网站516年结果。gydF4y2Ba3月Micropaleontol。gydF4y2Ba14gydF4y2Ba,119 - 152 (1989)。gydF4y2Ba
Gradstein, f M。,Ogg, j·G。,Schmitz, M. & Ogg, G. (eds)2012年地质时间尺度gydF4y2Ba(爱思唯尔,2012)。gydF4y2Ba
Florindo, f . et al。新的magnetobiostratigraphic年表和当前的变化在渐新世第三纪中新世边界DSDP网站516(格兰德河上升,西南大西洋)。gydF4y2Ba古海洋学gydF4y2Ba30.gydF4y2Ba,659 - 681 (2015)。gydF4y2Ba
布莱恩,w . b . &邓肯,r . a .年龄和碎屑的出处的视野从洞516 f。深海钻探计划的初步报告,72卷,475 - 477。gydF4y2Bahttps://doi.org/10.2973/dsdp.proc.72.118.1983gydF4y2Ba(美国政府印刷局,1983)。gydF4y2Ba
汉密尔顿:白垩纪/三级边界研究深海钻探计划网站516年格兰德河上升,南大西洋:合成。深海钻探计划的初步报告,72卷,949 - 952。gydF4y2Bahttps://doi.org/10.2973/dsdp.proc.72.150.1983gydF4y2Ba(美国政府印刷局,1983)。gydF4y2Ba
皮尔森,G。,我Premoli席尔瓦。,H一个ggerty, J. A. & Shipboard Scientific Party (PANGAEA, 2005).
皮尔森,p . n .浮游有孔虫生物地层学的发展远洋上限平顶海山在马绍尔群岛。《海洋钻井程序、科学的结果,卷144,21-59。(德州农工大学,1995)。gydF4y2Ba
Westerhold, T。Rohl U。,Donner, B. & Zachos, J. C. Global extent of early Eocene hyperthermal events: a new Pacific benthic foraminiferal isotope record from Shatsky Rise (ODP Site 1209).Paleoceanogr。Paleoclimatol。gydF4y2Ba33gydF4y2Ba,626 - 642 (2018)。gydF4y2Ba
马龙,m . J。,Bralower, t·J。,我Premoli席尔瓦。&Shipboard Scientific Party. Paleontological investigation on planktonic foraminifers of ODP Hole 144-872C (PANGAEA, 2005).
帕加尼,M。,一个rthur, M. A. & Freeman, K. H. Miocene evolution of atmospheric carbon dioxide.古海洋学gydF4y2Ba14gydF4y2Ba,273 - 292 (1999)。gydF4y2Ba
•鲁丁曼,w·F。,Sarnthein, M. & Shipboard Scientific Party. P-wave velocity (PWL whole-core system) on ODP Hole 108-664B (PANGAEA, 2005).
什么不对的p F。,Kristoffersen, Y. & Shipboard Scientific Party. P-wave velocity (PWS split-core system) on ODP Hole 114-698A (PANGAEA, 2005).
Leutert, t·J。Auderset,。,Martínez-Garci, A., Modestou, S. & Meckler, A. N. Coupled Southern Ocean cooling and Antarctic ice sheet expansion during the middle Miocene.Geosci Nat。gydF4y2Ba13gydF4y2Ba,634 - 639 (2020)。gydF4y2Ba
Sangiorgi F。,Quaijtaal, W., Donders, T. H., Schouten, S. & Louwye, S. Middle Miocene temperature and productivity evolution at a Northeast Atlantic shelf site (IODP U1318, Porcupine Basin): global and regional changes.Paleoceanogr。Paleoclimatol。gydF4y2Ba36gydF4y2Bae2020PA004059 (2021)。gydF4y2Ba
Prell, w . L。,N我我tsuma, N. & al, E. Site 730. Proceedings of the Ocean Drilling Program, Initial Reports, Vol. 117, 555–584.https://doi.org/10.2973/odp.proc.ir.117.117.1989gydF4y2Ba(德州农工大学,1989)。gydF4y2Ba
皮尔斯,j·W。,Weissel, J. K. & Shipboard Scientific Party. Hole/core summary of ODP Site 121-757. (PANGAEA, 2005).
内森,s a & Leckie r . m .中新世浮游有孔虫生物地层学的网站1143年和1146年,184年ODP的腿,南中国海。《海洋钻井程序、科学的结果,184卷,1。gydF4y2Bahttps://doi.org/10.2973/odp.proc.sr.184.219.2003gydF4y2Ba(德州农工大学,2003)。gydF4y2Ba
Censarek, b & Gersonde r数据报告:相对丰富和中新世地层范围选定的硅藻的部分在ODP网站689年,690年、1088年和1092年(南大洋的大西洋扇区)。《海洋钻井程序、科学的结果,177卷,1 - 14。gydF4y2Bahttps://doi.org/10.2973/odp.proc.sr.177.119.2003gydF4y2Ba(德州农工大学,2003)。gydF4y2Ba
论著,共和党和杰克逊,m . l .氧化铁从土壤和粘土去除dithionite-citrate系统与碳酸氢钠缓冲。gydF4y2Ba粘土的矿工。gydF4y2Ba7gydF4y2Ba,317 - 327 (1960)。gydF4y2Ba
徐文伯r s &亨德里克斯,美国纳克亚硝酸盐和硝酸盐测定钒在环境和生物材料与化学发光检测(3)减少。gydF4y2Ba肛交。化学。gydF4y2Ba61年gydF4y2Ba,2715 - 2718 (1989)。gydF4y2Ba
西格曼,d . et al .硝酸细菌的氮同位素分析方法在海水和淡水。gydF4y2Ba肛交。化学。gydF4y2Ba73年gydF4y2Ba,4145 - 4153 (2001)。gydF4y2Ba
Casciotti, k . L。西格曼,d . M。,H一个stings, M. G., Böhlke, J. & Hilkert, A. Measurement of the oxygen isotopic composition of nitrate in seawater and freshwater using the denitrifier method.肛交。化学。gydF4y2Ba74年gydF4y2Ba,4905 - 4912 (2002)。gydF4y2Ba
McIlvin m r & Casciotti k . l .技术更新硝酸细菌方法同位素分析。gydF4y2Ba肛交。化学。gydF4y2Ba83年gydF4y2Ba,1850 - 1856 (2011)。gydF4y2Ba
Weigand, m·A。Foriel, J。,Barnett, B., Oleynik, S. & Sigman, D. M. Updates to instrumentation and protocols for isotopic analysis of nitrate by the denitrifier method.快速Commun。质量范围。gydF4y2Ba30.gydF4y2Ba,1365 - 1383 (2016)。gydF4y2Ba
Straub, m . et al .北大西洋固氮作用的变化由海洋环流控制。gydF4y2Ba自然gydF4y2Ba501年gydF4y2Ba,200 - 203 (2013)。gydF4y2Ba
Martinez-Garcia, a . et al .铁质肥料亚南极的海洋在最后一个冰河时代。gydF4y2Ba科学gydF4y2Ba343年gydF4y2Ba,1347 - 1350 (2014)。gydF4y2Ba
Leichliter, j . n . et al .氮同位素在牙釉质记录饮食和营养级浓缩:控制饲养试验结果。gydF4y2Ba化学。青烟。gydF4y2Ba563年gydF4y2Ba120047 (2021)。gydF4y2Ba
Auderset,。,Schmitt, M. & Martínez-García, A. Simultaneous extraction and chromatographic separation ofngydF4y2Ba烷烃和alkenones甘油二烷基甘油四醚通过选择性加速溶剂萃取。gydF4y2BaOrg。Geochem。gydF4y2Ba143年gydF4y2Ba103979 (2020)。gydF4y2Ba
Patwardhan, a p & Thompson d·h·高效合成买票其他48四醚大环的bisphosphocholines。gydF4y2Ba有机的信gydF4y2Ba1gydF4y2Ba,241 - 244 (1999)。gydF4y2Ba
Hopmans, e . C。思,S。&Damsté, J. S. S. The effect of improved chromatography on GDGT-based palaeoproxies.Org。Geochem。gydF4y2Ba93年gydF4y2Ba1 - 6 (2016)。gydF4y2Ba
聪明,s . m . et al .脚踏实地planktic foraminifer-bound氮同位素paleo-proxy马尾藻海。gydF4y2BaGeochim。Cosmochim。学报gydF4y2Ba235年gydF4y2Ba,463 - 482 (2018)。gydF4y2Ba
&聪明,s m . et al .氮同位素组成的组织和壳牌绑定有机物等planktic有孔虫在南大洋表面水域。gydF4y2BaGeochem。地球物理学。呈gydF4y2Ba21gydF4y2Bae2019GC008440 (2020)。gydF4y2Ba
Ai, x大肠et al。南太平洋上升流,地球的倾斜,glacial-interglacial大气COgydF4y2Ba2gydF4y2Ba改变。gydF4y2Ba科学gydF4y2Ba370年gydF4y2Ba,1348 - 1352 (2020)。gydF4y2Ba
Auderset, a . et al .墨西哥湾流强化上新世早期中美洲航道的变浅。gydF4y2Ba地球的星球。科学。列托人。gydF4y2Ba520年gydF4y2Ba,268 - 278 (2019)。gydF4y2Ba
场,p . k . et al .早期早第三纪的温度西南太平洋的进化。gydF4y2Ba自然gydF4y2Ba461年gydF4y2Ba,776 - 779 (2009)。gydF4y2Ba
场,p . k . et al。始新世早期冷却与流在塔斯马尼亚网关。gydF4y2BaProc。《科学。gydF4y2Ba110年gydF4y2Ba,9645 - 9650 (2013)。gydF4y2Ba
Boscolo Galazzo, f . et al。始新世中期气候适宜期(MECO): multiproxy记录古海洋东南大西洋的变化(ODP网站1263年,沃尔维斯岭)。gydF4y2Ba古海洋学gydF4y2Ba29日gydF4y2Ba,1143 - 1161 (2014)。gydF4y2Ba
伯吉斯,c, e . et al。始新世中期在南太平洋气候旋回性:对全球冰量的影响。gydF4y2Ba地质gydF4y2Ba36gydF4y2Ba,651 - 654 (2008)。gydF4y2Ba
Caley, t . et al .高纬度倾角作为主导迫使阿古拉斯海流系统。gydF4y2Ba爬。过去的gydF4y2Ba7gydF4y2Ba,1285 - 1296 (2011)。gydF4y2Ba
Cramwinckel, m . j . et al .温暖的分层,并限制拉布拉多海在始新世中期及其气候适宜期。gydF4y2BaPaleoceanogr。Paleoclimatol。gydF4y2Ba35gydF4y2Bae2020PA003932 (2020)。gydF4y2Ba
Cramwinckel, m . j . et al .同步热带和极地温度在始新世的进化。gydF4y2Ba自然gydF4y2Ba559年gydF4y2Ba,382 - 386 (2018)。gydF4y2Ba
德酒吧,m . W。,Rampen, S. W., Hopmans, E. C., Damsté, J. S. S. & Schouten, S. Constraining the applicability of organic paleotemperature proxies for the last 90 Myrs.Org。Geochem。gydF4y2Ba128年gydF4y2Ba,122 - 136 (2019)。gydF4y2Ba
De Vleeschouwer D。,Petrick, B. F. & Martínez‐García, A. Stepwise weakening of the pliocene Leeuwin current.地球物理学。卷。gydF4y2Ba46gydF4y2Ba,8310 - 8319 (2019)。gydF4y2Ba
道格拉斯,p . m . et al .明显的纬向异质性在始新世南部高纬度海洋表面温度。gydF4y2BaProc。《科学。gydF4y2Ba111年gydF4y2Ba,6582 - 6587 (2014)。gydF4y2Ba
香浓,j . et al .极端的温暖和heat-stressed浮游生物在热带地区Paleocene-Eocene热最大。gydF4y2Ba科学。睡觉。gydF4y2Ba3gydF4y2Bae1600891 (2017)。gydF4y2Ba
香浓,j . & Sluijs a向量化环境重建从古代non-analogue微体化石组合:生态Paleocene-Eocene甲藻的偏好。gydF4y2BaEarth-Sci。牧师。gydF4y2Ba185年gydF4y2Ba,956 - 973 (2018)。gydF4y2Ba
水平,j . et al .中间纬度温度变化在渐新世到中新世早期。gydF4y2BaPaleoceanogr。Paleoclimatol。gydF4y2Ba34gydF4y2Ba,1328 - 1343 (2019)。gydF4y2Ba
哈特曼,j . D。,Sangiorgi F。&Escutia, C. Paleoceanography and ice sheet variability offshore Wilkes Land, Antarctica – Part 3: insights from Oligocene–Miocene TEX86年gydF4y2Ba的海洋表面温度重建。gydF4y2Ba爬。过去。gydF4y2Ba14gydF4y2Ba,1275 - 1297 (2018)。gydF4y2Ba
海耶斯,c . t . et al。在南大西洋深处一个停滞的事件在过去的间冰期。gydF4y2Ba科学gydF4y2Ba346年gydF4y2Ba,1514 - 1517 (2014)。gydF4y2Ba
霍利斯,c . j . et al .热带海洋温度在高纬度地区在始新世南太平洋。gydF4y2Ba地质gydF4y2Ba37gydF4y2Ba,99 - 102 (2009)。gydF4y2Ba
霍利斯,c . j . et al .早第三纪早期西南太平洋的温度历史:协调代理和模型。gydF4y2Ba地球的星球。科学。列托人。gydF4y2Ba349年gydF4y2Ba53 - 66 (2012)。gydF4y2Ba
Inglis, g . n . et al .后裔向冰室:始新世海洋表面冷却从GDGT分布推断。gydF4y2Ba古海洋学gydF4y2Ba30.gydF4y2Ba,1000 - 1020 (2015)。gydF4y2Ba
Keating-Bitonti, c R。、Ivany l . C。、Affek h P。,Douglas, P. & Samson, S. D. Warm, not super-hot, temperatures in the early Eocene subtropics.地质gydF4y2Ba39gydF4y2Ba,771 - 774 (2011)。gydF4y2Ba
劳伦斯,k . et al .比较新第三纪末UgydF4y2Bak′gydF4y2Ba37gydF4y2Ba和泰克斯gydF4y2Ba86年gydF4y2Ba从赤道东太平洋古温度记录轨道决议。gydF4y2BaPaleoceanogr。Paleoclimatol。gydF4y2Ba35gydF4y2Bae2020PA003858 (2020)。gydF4y2Ba
Leutert, t·J。Auderset,。,Martínez-García, A., Modestou, S. & Meckler, A. N. Coupled Southern Ocean cooling and Antarctic ice sheet expansion during the middle Miocene.Geosci Nat。gydF4y2Ba13gydF4y2Ba,634 - 639 (2020)。gydF4y2Ba
刘,z . et al . Global cooling Eocene-Oligocene气候过渡。gydF4y2Ba科学gydF4y2Ba323年gydF4y2Ba,1187 - 1190 (2009)。gydF4y2Ba
Naafs, b . d . A。Voelker,。卡拉斯,C。,一个ndersen, N. & Sierro, F. Repeated near‐collapse of the Pliocene sea surface temperature gradient in the North Atlantic.Paleoceanogr。Paleoclimatol。gydF4y2Ba35gydF4y2Bae2020PA003905 (2020)。gydF4y2Ba
O ' brien, c . l . et al。高在热带温暖的海洋表面温度池在上新世。gydF4y2BaGeosci Nat。gydF4y2Ba7gydF4y2Ba,606 - 611 (2014)。gydF4y2Ba
O ' brien, c . l . et al。渐新世气候和全球表面温度进化的谜。gydF4y2BaProc。《科学。gydF4y2Ba117年gydF4y2Ba,25302 - 25309 (2020)。gydF4y2Ba
皮尔森,p . n . et al .稳定通过始新世温暖的热带气候。gydF4y2Ba地质gydF4y2Ba35gydF4y2Ba,211 - 214 (2007)。gydF4y2Ba
Petrick, b . et al。冰川印尼通流削弱整个mid-Pleistocene气候过渡。gydF4y2Ba科学。代表。gydF4y2Ba9gydF4y2Ba16995 (2019)。gydF4y2Ba
Petrick, b . et al。海洋和气候演变的东南部亚热带大西洋过去3.5 Ma。gydF4y2Ba地球的星球。科学。列托人。gydF4y2Ba492年gydF4y2Ba12-21 (2018)。gydF4y2Ba
Rommerskirchen F。康登,T。,Mollenhauer, G., Dupont, L. & Schefus, E. Miocene to Pliocene development of surface and subsurface temperatures in the Benguela Current system.古海洋学gydF4y2Ba26gydF4y2BaPA3216 (2011)。gydF4y2Ba
Sangiorgi, f . et al。海洋变暖南部和威尔克斯地在mid-Miocene冰原撤退。gydF4y2BaCommun Nat。gydF4y2Ba9gydF4y2Ba317 (2018)。gydF4y2Ba
Śliwińska K·K。Thomsen E。,思,S。,Schoon, P. L. & Heilmann-Clausen, C. Climate- and gateway-driven cooling of Late Eocene to earliest Oligocene sea surface temperatures in the North Sea Basin.科学。代表。gydF4y2Ba9gydF4y2Ba4458 (2019)。gydF4y2Ba
Sluijs, a . et al .亚热带北极海洋温度在古新世、始新世热最大。gydF4y2Ba自然gydF4y2Ba441年gydF4y2Ba,610 - 613 (2006)。gydF4y2Ba
Sluijs, a . et al . Paleocene-early始新世末北冰洋海面温度:重新评估生物标志物在罗蒙诺索夫海岭古温度测定。gydF4y2Ba爬。过去的gydF4y2Ba16gydF4y2Ba,2381 - 2400 (2020)。gydF4y2Ba
Sluijs et al。环境前体快速光碳注入在古新世、始新世边界。gydF4y2Ba自然gydF4y2Ba450年gydF4y2Ba,1218 - 1221 (2007)。gydF4y2Ba
史密斯,r . a . et al . Plio更新世应承担的印尼直流变化推动东部印度洋海表温度。gydF4y2BaPaleoceanogr。Paleoclimatol。gydF4y2Ba35gydF4y2Bae2020PA003872 (2020)。gydF4y2Ba
斯托克,e·W。,Jones, M. T., Tierney, J. E., Svensen, H. H. & Whiteside, J. H. Temperature changes across the Paleocene-Eocene Thermal Maximum – a new high-resolution TEX86年gydF4y2Ba从北海盆地东部温度记录。gydF4y2Ba地球的星球。科学。列托人。gydF4y2Ba544年gydF4y2Ba116388 (2020)。gydF4y2Ba
超级,j . r . et al .温度和pCO北大西洋gydF4y2Ba2gydF4y2Ba耦合在早中中新世。gydF4y2Ba地质gydF4y2Ba46gydF4y2Ba,519 - 522 (2018)。gydF4y2Ba
维氏,m . l . et al .寒冷北欧海域法术在始新世早期温室。gydF4y2BaCommun Nat。gydF4y2Ba11gydF4y2Ba4713 (2020)。gydF4y2Ba
韦德,b . s . et al . Multiproxy突然海面的记录冷却整个Eocene-Oligocene墨西哥湾的过渡。gydF4y2Ba地质gydF4y2Ba40gydF4y2Ba,159 - 162 (2012)。gydF4y2Ba
Wittkopp, f .有机沉积物的地球化学调查日本海上:跟踪以来古海洋、古气候变化mid-Miocene(泛大陆,2017)。gydF4y2Ba
Zachos, j . c . et al .极端气候变暖的中纬度地区沿海海洋Paleocene-Eocene热最大:从特克斯推论gydF4y2Ba86年gydF4y2Ba和同位素数据。gydF4y2Ba地质gydF4y2Ba34gydF4y2Ba,737 - 740 (2006)。gydF4y2Ba
张,y G。帕加尼,M。刘,z . H。,Bohaty, S. M. & DeConto, R. A 40-million-year history of atmospheric CO2gydF4y2Ba。gydF4y2Ba费罗斯。反式。r . Soc。一个数学。理论物理。Eng。科学。gydF4y2Ba371年gydF4y2Ba,20130096 (2013)。gydF4y2Ba
张,y G。帕加尼,M。&Liu, Z. A 12-million-year temperature history of the tropical Pacific Ocean.科学gydF4y2Ba344年gydF4y2Ba,84 - 87 (2014)。gydF4y2Ba
壮族,G。,帕加尼,M。&张,y G。Monsoonal upwelling in the western Arabian Sea since the middle Miocene.地质gydF4y2Ba45gydF4y2Ba,655 - 658 (2017)。gydF4y2Ba
金,黄永发。et al。新指数和校准源自crenarchaeal分布的类异戊二烯四醚脂质:对过去的海洋表面温度重建的影响。gydF4y2BaGeochim。Cosmochim。学报gydF4y2Ba74年gydF4y2Ba,4639 - 4654 (2010)。gydF4y2Ba
Tierney, j . e . & Tingley m·p·贝叶斯空间变化特克斯校准模型gydF4y2Ba86年gydF4y2Ba代理。gydF4y2BaGeochim。Cosmochim。学报gydF4y2Ba127年gydF4y2Ba,83 - 106 (2014)。gydF4y2Ba
Weijers, j·w·H。思,S。,Spaargaren, O. C. & Sinninghe Damsté, J. S. Occurrence and distribution of tetraether membrane lipids in soils: implications for the use of the TEX86年gydF4y2Ba代理和指数。gydF4y2BaOrg。Geochem。gydF4y2Ba37gydF4y2Ba,1680 - 1693 (2006)。gydF4y2Ba
张,y . g . et al .甲烷指数:四醚热点脂质生物标志物指标检测海洋天然气水合物的稳定。gydF4y2Ba地球的星球。科学。列托人。gydF4y2Ba307年gydF4y2Ba,525 - 534 (2011)。gydF4y2Ba
张,y G。帕加尼,M。&Wang, Z. Ring Index: a new strategy to evaluate the integrity of TEX86年gydF4y2Ba古温度测定。gydF4y2Ba古海洋学gydF4y2Ba31日gydF4y2Ba,220 - 232 (2016)。gydF4y2Ba
Hopmans, e . c . et al。小说代表陆地沉积物中有机质基于分支和类异戊二烯四醚脂质。gydF4y2Ba地球的星球。科学。列托人。gydF4y2Ba224年gydF4y2Ba,107 - 116 (2004)。gydF4y2Ba
Peterse F。,N我col, G. W., Schouten, S. & Damsté, J. S. S. Influence of soil pH on the abundance and distribution of core and intact polar lipid-derived branched GDGTs in soil.Org。Geochem。gydF4y2Ba41gydF4y2Ba,1171 - 1175 (2010)。gydF4y2Ba
Fietz, S。,Martínez‐Garcia, A., Huguet, C., Rueda, G. & Rosell‐Melé, A. Constraints in the application of the Branched and Isoprenoid Tetraether index as a terrestrial input proxy.j .地球物理学。海洋》gydF4y2Ba116年gydF4y2BaC10032 (2011)。gydF4y2Ba
Fietz, s . et al。共变crenarchaeol和分支GDGTs在全球分布的海洋和淡水沉积档案。gydF4y2Ba水珠。星球。改变gydF4y2Ba92年gydF4y2Ba,275 - 285 (2012)。gydF4y2Ba
·德容,c . et al .剧烈变化的分布支四醚脂质在悬浮物和沉积物从叶尼塞河和喀拉海(西伯利亚):对使用brGDGT-based代理在沿海海洋沉积物。gydF4y2BaGeochim。Cosmochim。学报gydF4y2Ba165年gydF4y2Ba,200 - 225 (2015)。gydF4y2Ba
肖,w . et al .无处不在生产支甘油二烷基甘油四醚(brGDGTs)全球海洋环境:一个新的brGDGTs指标来源。gydF4y2BaBiogeosciencesgydF4y2Ba13gydF4y2Ba,5883 - 5894 (2016)。gydF4y2Ba
女人,c . et al .选择性保护土壤有机质的氧化海洋沉积物(马德拉深海平原)。gydF4y2BaGeochim。Cosmochim。学报gydF4y2Ba72年gydF4y2Ba,6061 - 6068 (2008)。gydF4y2Ba
Damste j·s·s .的空间异质性的来源支四醚在货架系统:地球化学的四醚伯劳河三角洲(加里曼丹,印度尼西亚)。gydF4y2BaGeochim。Cosmochim。学报gydF4y2Ba186年gydF4y2Ba13-31 (2016)。gydF4y2Ba
确认gydF4y2Ba
这项工作是由马克斯·普朗克协会(MPG), SNSF流动性格兰特P2EZP2_200000和塔特尔基金系的地质,普林斯顿大学。沉积物样品为本研究提供的国际海洋探索项目。我们承认m·施密特f . Rubach b Hinnenberg和美国Neugebauer实验室协助。我们也感谢n .沙编程的支持。我们感谢c·多伊奇的讨论。gydF4y2Ba
资金gydF4y2Ba
开放获取的资金由马克斯·普朗克协会提供。gydF4y2Ba
作者信息gydF4y2Ba
作者和联系gydF4y2Ba
贡献gydF4y2Ba
嗜和A.M.-G。设计研究。嗜测量foraminifer-bound氮同位素和有机生物标志物A.M.-G的实验室。嗜,D.M.S. and A.M.-G. wrote the manuscript. B.T. and P.-R.E. contributed to sample preparation and foraminifer ecology, supervised by R.S. All authors contributed to the interpretation of the data and provided input to the final manuscript at different stages of the project.
相应的作者gydF4y2Ba
道德声明gydF4y2Ba
相互竞争的利益gydF4y2Ba
作者宣称没有利益冲突。gydF4y2Ba
同行评审gydF4y2Ba
同行审查的信息gydF4y2Ba
自然gydF4y2Ba谢谢芭贝特Hoogakker Bernhard Naafs,丽贝卡·罗宾逊和其他,匿名的,审稿人(s)为他们的贡献的同行评审工作。gydF4y2Ba
额外的信息gydF4y2Ba
出版商的注意gydF4y2Ba施普林格自然保持中立在发表关于司法主权地图和所属机构。gydF4y2Ba
扩展数据数据和表gydF4y2Ba
扩展数据图1新生代foraminifer-bound氮同位素和含氮量数据编译、网站和分类单元表示。gydF4y2Ba
Foraminifer-bound N内容(gydF4y2Ba一个gydF4y2Ba)和FB-δgydF4y2Ba15gydF4y2BaN (gydF4y2BabgydF4y2Ba)516年DSDP网站特有的分析,ODP网站872年和1211年ODP网站,辅以数据混合分类单元和具体属清晨et al。gydF4y2Ba18gydF4y2Ba1263年和1209年对ODP网站。我们编译的推定地共生类群或粗分数由共生类群,避免需要正确的共生和symbiont-barren物种之间的典型的抵消gydF4y2Ba19gydF4y2Ba,gydF4y2Ba89年gydF4y2Ba。交会图法之间的δgydF4y2Ba15gydF4y2BaN和N含量没有相关性。gydF4y2Ba
扩展数据图2新生代特克斯gydF4y2Ba86年BayspargydF4y2Ba派生的SST编译。gydF4y2Ba
太平洋分类中低纬度地区过渡和上升流区域(gydF4y2Ba一个gydF4y2Ba)和高纬度地区(gydF4y2BabgydF4y2Ba)。灰色行表示底栖生物氧同位素编译Westerhold et al。gydF4y2Ba8gydF4y2Ba。gydF4y2BacgydF4y2Ba与核心位置,地图:现代地图显示当前位置的核心网站。Palaeomap 20 Ma palaeorecords跨越10 ~ 30 Ma和Palaeomap与马palaeorecords跨越40 - 50 Ma。地图是改编自海洋钻井地层网络(ODSN) Paleomap项目(gydF4y2Bahttp://www.odsn.de/odsn/services/paleomap/paleomap.htmlgydF4y2Ba,2022年5月)被访问。古纬度为核心网站后计算货车Hinsbergen et al。gydF4y2Ba52gydF4y2Ba。gydF4y2Ba
扩展数据图3新生代特克斯gydF4y2Ba86年BayspargydF4y2Ba派生的SST编译、分类的基础上自由。gydF4y2Ba
一个gydF4y2Ba德克萨斯州gydF4y2Ba86年BayspargydF4y2Ba海温资料导出生成的在这个研究是用红色(DSDP网站516),粉红色(DSDP网站588),暗橙色(ODP网站667),蓝色(ODP网站704),亮绿色(ODP网站730),绿色(ODP网站754),紫色光(ODP网站1146)和橙色(ODP网站1263)。在灰色的海温记录发表我们的编译(补充数据gydF4y2Ba2gydF4y2Ba)。开放点的不同颜色显示样本排除由于异常(见GDGTs分布gydF4y2Ba方法gydF4y2Ba)。gydF4y2BabgydF4y2Ba德克萨斯州gydF4y2Ba86年BayspargydF4y2Ba派生的SST记录分类在低(0-20°N / S,红色)和中等(20 - 40°N / S,橙色)纬度,过渡区域(40 - 50°N / S,浅蓝色),高纬度地区(> 50°N / S,蓝色)和上升流区域(绿色)。黑线表示底栖生物氧同位素编译gydF4y2Ba8gydF4y2Ba。gydF4y2Ba
扩展数据图4比较特克斯和置信区间gydF4y2Ba86年gydF4y2Ba派生的SST重建。gydF4y2Ba
一个gydF4y2Ba德克萨斯州gydF4y2Ba86年BayspargydF4y2Ba派生的海温为中、低(橙色)和高(蓝色)纬度数据与相应的90%置信区间。gydF4y2BabgydF4y2Ba,比较δgydF4y2Ba15gydF4y2BaN-gradient(ΔδgydF4y2Ba15gydF4y2BaNgydF4y2Ba便士gydF4y2Ba)和纬向SST-gradient(ΔSSTgydF4y2BaL-HgydF4y2Ba)计算,使用两个风场校准:风场gydF4y2BaBayspargydF4y2BaTierney和Tingley之后gydF4y2Ba138年gydF4y2Ba(90%置信区间传播gydF4y2Ba一个gydF4y2Ba)和海温gydF4y2BaTEXHgydF4y2Ba之后金等。gydF4y2Ba137年gydF4y2Ba。gydF4y2BacgydF4y2Ba纬度的特克斯gydF4y2Ba86年gydF4y2Ba梯度(ΔTEXgydF4y2Ba86年L-HgydF4y2Ba)相比,提出的纬向风场梯度Cramwinckel et al。gydF4y2Ba99年gydF4y2Ba。(ΔgydF4y2BaTgydF4y2Ba= SSTgydF4y2BaTEXH86gydF4y2Ba−SSTgydF4y2Baδ18OgydF4y2Ba,黑色线)。请注意,ΔTEXgydF4y2Ba86年L-HgydF4y2Ba不依赖于所使用的校准。gydF4y2Ba
扩展数据模型与图5 GTS12-adjusted年龄年龄领带点。gydF4y2Ba
一个gydF4y2Ba516年,DSDP网站(参考文献。gydF4y2Ba57gydF4y2Ba,gydF4y2Ba58gydF4y2Ba,gydF4y2Ba59gydF4y2Ba,gydF4y2Ba61年gydF4y2Ba,gydF4y2Ba62年gydF4y2Ba,gydF4y2Ba63年gydF4y2Ba)。gydF4y2BabgydF4y2Ba588年,DSDP网站(ref。gydF4y2Ba68年gydF4y2Ba)。gydF4y2BacgydF4y2Ba667年,ODP网站(ref。gydF4y2Ba69年gydF4y2Ba)。gydF4y2BadgydF4y2Ba690年,ODP网站(ref。gydF4y2Ba41gydF4y2Ba)。gydF4y2BaegydF4y2Ba704年,ODP网站(ref。gydF4y2Ba70年gydF4y2Ba)。gydF4y2BafgydF4y2Ba730年,ODP网站(ref。gydF4y2Ba73年gydF4y2Ba)。gydF4y2BaggydF4y2Ba738年,ODP网站(ref。gydF4y2Ba41gydF4y2Ba)。gydF4y2BahgydF4y2Ba754年,ODP网站(ref。gydF4y2Ba74年gydF4y2Ba)。gydF4y2Ba我gydF4y2Ba872年,ODP网站(ref。gydF4y2Ba65年gydF4y2Ba)。gydF4y2BajgydF4y2Ba1146年,ODP网站(ref。gydF4y2Ba75年gydF4y2Ba)。gydF4y2BakgydF4y2Ba1209年,ODP网站(参考文献。gydF4y2Ba18gydF4y2Ba,gydF4y2Ba66年gydF4y2Ba)。gydF4y2BalgydF4y2Ba1211年,ODP网站(ref。gydF4y2Ba67年gydF4y2Ba)。gydF4y2Ba米gydF4y2Ba1263年,ODP网站(参考文献。gydF4y2Ba8gydF4y2Ba,gydF4y2Ba18gydF4y2Ba)。gydF4y2Ba
补充信息gydF4y2Ba
补充数据1gydF4y2Ba
FB-δ15N和FB-N内容数据。表题为《DSDP 516”、“ODP 872”包含所有物种特异性,genus-specific和mixed-taxa foraminifer-boundδ15N和FB-N内容从DSDP测量网站516年和872年ODP网站,分别绘制在无花果。gydF4y2Ba2gydF4y2Ba和gydF4y2Ba3gydF4y2Ba和扩展数据图。gydF4y2Ba1gydF4y2Ba。表题为“ODP 1211”包含mixed-taxa FB-δ15N和FB-N内容测量从ODP网站1211年,扩展数据图的绘制。gydF4y2Ba1gydF4y2Ba。表题为《ODP 1209’和‘ODP 1263包含genus-specific和mixed-taxa FB-δ15N和FB-N内容测量从ODP网站1209年和1263年,分别测量了裁判。gydF4y2Ba18gydF4y2Ba修改年龄模型,绘制在无花果。gydF4y2Ba2gydF4y2Ba和gydF4y2Ba3gydF4y2Ba和扩展数据图。gydF4y2Ba1gydF4y2Ba。表题为“d15N梯度”包含氮同位素太平洋和大西洋之间的梯度计算绘制在无花果。gydF4y2Ba2gydF4y2Ba和gydF4y2Ba3gydF4y2Ba和扩展数据图。gydF4y2Ba4gydF4y2Ba。gydF4y2Ba
补充数据2gydF4y2Ba
特克斯gydF4y2Ba86年gydF4y2Ba数据以本研究(ODP / DSDP网站516,588,667,704,730,754,1146和1263)和编译所有的海洋特克斯出版gydF4y2Ba86年gydF4y2Ba记录。这个表包含了所有GDGTs测量,计算指标,温度和梯度。gydF4y2Ba
补充数据3gydF4y2Ba
年龄模型申请ODP / DSDP网站516,588,667,690,704,730,738,754,1146,1209年、1211年和1263年与生物地层的(钙质微型浮游生物和有孔虫)和磁性地层论据调整GTS12时间表。gydF4y2Ba
权利和权限gydF4y2Ba
开放获取gydF4y2Ba本文是基于知识共享署名4.0国际许可,允许使用、共享、适应、分布和繁殖在任何媒介或格式,只要你给予适当的信贷原始作者(年代)和来源,提供一个链接到Creative Commons许可,并指出如果变化。本文中的图片或其他第三方材料都包含在本文的创作共用许可,除非另有说明在一个信用额度的材料。如果材料不包括在本文的创作共用许可证和用途是不允许按法定规定或超过允许的使用,您将需要获得直接从版权所有者的许可。查看本许可证的副本,访问gydF4y2Bahttp://creativecommons.org/licenses/by/4.0/gydF4y2Ba。gydF4y2Ba
关于这篇文章gydF4y2Ba

引用这篇文章gydF4y2Ba
Auderset,。,Moretti, S., Taphorn, B.et al。gydF4y2Ba加强海洋氧化新生代温暖的时期。gydF4y2Ba自然gydF4y2Ba609年gydF4y2Ba,77 - 82 (2022)。https://doi.org/10.1038/s41586 - 022 - 05017 - 0gydF4y2Ba
收到了gydF4y2Ba:gydF4y2Ba
接受gydF4y2Ba:gydF4y2Ba
发表gydF4y2Ba:gydF4y2Ba
发行日期gydF4y2Ba:gydF4y2Ba
DOIgydF4y2Ba:gydF4y2Bahttps://doi.org/10.1038/s41586 - 022 - 05017 - 0gydF4y2Ba
评论gydF4y2Ba
通过提交评论你同意遵守我们的gydF4y2Ba条款gydF4y2Ba和gydF4y2Ba社区指导原则gydF4y2Ba。如果你发现一些滥用或不符合我们的条件或准则请国旗是不合适的。gydF4y2Ba
