








文摘gydF4y2Ba
原始热带森林被广泛公认为是非常重要的生物多样性和较高的生物量gydF4y2Ba1gydF4y2Ba。相反,砍伐热带森林通常描述为退化的生态系统gydF4y2Ba2gydF4y2Ba。然而,是否记录结果在退化生态系统功能不太清楚:关键生态过程的强度和韧性的变化在大套房的物种很少在生态综合和量化评估框架。这里我们采用一个生态系统能量镜头获得新的见解热带森林干扰的影响关键生态功能的整合方面:食物途径和鸟类和哺乳动物的群落结构。我们专注于一个梯度生成原始并记录在婆罗洲森林和油棕种植园。在被砍伐的森林资源消耗总量增加2.5倍,鸟类和哺乳动物相比,在原始森林,可能受到更大的资源可访问性和植被适口性。最主要的精力充沛的通路保持较高的物种多样性和冗余,这意味着保持弹性。被砍伐的森林转化为油棕种植园导致的崩溃最充满活力的途径。远非退化生态系统,甚至大量砍伐森林可以用增强活力和多样的生态系统的生态功能水平。gydF4y2Ba
主要gydF4y2Ba
人造森林,如选择性砍伐森林,经常被描绘成退化生态系统,因为他们改变结构和生物量较低。生态系统退化的概念可以是一把双刃剑。它正确地把人的注意力吸引到了保护古老的价值系统和生态系统恢复的重要性。然而,它也可以表明人造生态系统是生态价值较低,因此,在某些情况下,适合农业转换(如油棕种植园)和其他土地使用gydF4y2Ba3gydF4y2Ba,gydF4y2Ba4gydF4y2Ba,gydF4y2Ba5gydF4y2Ba。gydF4y2Ba
择伐和其他形式的结构性改变森林越来越盛行的植被的热带森林生态区gydF4y2Ba2gydF4y2Ba。这种干扰频繁导致原始专家物种下降gydF4y2Ba1gydF4y2Ba在某些情况下,还在非专业物种gydF4y2Ba6gydF4y2Ba,gydF4y2Ba7gydF4y2Ba,gydF4y2Ba8gydF4y2Ba。然而,species-focused生物多样性指标只有一个衡量生态系统的活力和功能,而且很少考虑物种的集体作用套件在维护生态功能gydF4y2Ba9gydF4y2Ba。gydF4y2Ba
另一种方法是把重点放在关键分类群的能量,和的数量和相对优势物种造成每一个充满活力的途径。精力充沛的方法来研究生态系统结构和功能在生态系统有很长的历史gydF4y2Ba10gydF4y2Ba。几乎所有的生态系统是由一连串的捕获阳光通过数组的自养生物组织和分层组合的食草动物,食肉动物和腐食者。精力充沛的方法在相对意义上发光能量流动的关键类群,并提供洞察形状生物多样性和生态系统功能的过程。共同货币的能量使不同的公会和类群比较统一,身体上有意义的方式:主要精力充沛的通路可以被识别,每个通路的韧性,可以评估个别物种的丧失。定量之间的链接可以让动物社区和植物生态系统生产力他们所依赖。充满活力的途径尤其是动物群体的大小往往是关键指标相关的生态系统过程,如养分循环、种子散布和授粉,或营养因素如捕食压力的强度或可用性的资源供应,所有常见的度量下的统一能量通量gydF4y2Ba11gydF4y2Ba,gydF4y2Ba12gydF4y2Ba。gydF4y2Ba
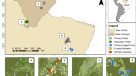
能量方法很少被应用在热带生态系统生物多样性,因为他们需要观察的范围gydF4y2Ba11gydF4y2Ba,gydF4y2Ba12gydF4y2Ba,gydF4y2Ba13gydF4y2Ba。这样的分析依赖于:人口密度估计大量的物种;理解饮食和觅食行为的物种;和净初级生产力(NPP)的可靠估计。这里我们利用独特丰富的数据集应用能量镜头检测和量化方面的生态功能和活力的栖息地在沙巴,马来西亚,组成原始森林,砍伐森林和油棕种植园(无花果。gydF4y2Ba1gydF4y2Ba和扩展数据图。gydF4y2Ba1gydF4y2Ba)。我们的方法是计算食物能量的短期均衡生产或消费利率由特定物种,公会或分类组。我们专注于三个分类群(植物、鸟类和哺乳动物)常用的生物多样性和相对好理解生态指标。gydF4y2Ba

一个gydF4y2Ba- - - - - -gydF4y2BadgydF4y2Ba、地图显示的位置NPP情节和生物多样性调查在原始森林,砍伐森林和油棕种植园改变了森林生态系统的稳定性项目景观(gydF4y2Ba一个gydF4y2Ba),Maliau盆地(gydF4y2BabgydF4y2Ba),丹浓谷(gydF4y2BacgydF4y2Ba)和Sepilok (gydF4y2BadgydF4y2Ba)。的插图gydF4y2Ba一个gydF4y2Ba显示四个网站在沙巴的位置。绿色的阴影表明原始(深绿色)twice-logged(中间绿色)或严重记录(浅绿色)森林。相机和陷阱网格包括相机和小型哺乳动物陷阱。白色区域表明油棕种植园。gydF4y2Ba
我们感兴趣的是初级生产力的一部分被鸟类和哺乳动物,以及它如何沿着扰动梯度变化,和如何以及为什么各种食物能量通路在哺乳动物和鸟类,和物种的多样性导致这些路径,沿着扰动梯度变化。估计的104哺乳动物和144种鸟类密度三种生境类型,我们聚合数据从882年相机取样位置(42877相机陷阱的晚上),508年鸟点估算的位置,1488年小型陆地哺乳动物陷阱的位置(34058活动陷阱的夜晚)和336年蝙蝠陷阱的位置(图。gydF4y2Ba1gydF4y2Ba和扩展数据图。gydF4y2Ba1gydF4y2Ba)。然后我们每天精力充沛的支出计算每个物种基于他们的体重,分配每个物种一个饮食组和食品消费总额计算能量单位。对于初级生产力,我们依靠34 plot-years(情节的总和乘以每个情节的年数)监测的测量的关键部件NPP(树冠litterfall,伍迪增长,细根生产)使用全球生态系统监测网络的协议gydF4y2Ba14gydF4y2Ba,gydF4y2Ba15gydF4y2Ba,gydF4y2Ba16gydF4y2Ba在原始(gydF4y2BangydF4y2Ba= 4),记录(gydF4y2BangydF4y2Ba= 5)和油棕(gydF4y2BangydF4y2Ba= 1)的情节。这个数据集包含了超过14000的测量litterfall, 20000年树和罚款2700根直径测量样本。gydF4y2Ba
鸟类物种多样性总体保持在扰动梯度和山峰被砍伐的森林;对于哺乳动物,砍伐森林也略有增加,其次是油棕快速下降(无花果。gydF4y2Ba2 b, cgydF4y2Ba)。引人注目的是,鸟类和哺乳动物生物量显著增加(分别为144%和231%)记录森林原始森林相比,与哺乳动物贡献约75%的总生物量(鸟和哺乳动物)在两种生境类型(无花果。gydF4y2Ba2 b, cgydF4y2Ba)。gydF4y2Ba

一个gydF4y2Ba,总NPP沿着梯度(意味着集约1公顷的情节;gydF4y2BangydF4y2Ba= 4老增长(OG),gydF4y2BangydF4y2Ba为记录和= 5gydF4y2BangydF4y2Ba= 1为油棕(OP);误差是95%置信区间由单独测量的不确定性传播NPP组件),与单个数据点覆盖。gydF4y2BabgydF4y2Ba,gydF4y2BacgydF4y2Ba,总身体质量(酒吧、左轴)和物种的数量统计(蓝色的点和线,右轴)的鸟类(gydF4y2BabgydF4y2Ba)和哺乳动物(gydF4y2BacgydF4y2Ba)。gydF4y2BadgydF4y2Ba,gydF4y2BaegydF4y2Ba,总直接精力充沛的食物摄入量鸟类(gydF4y2BadgydF4y2Ba)和哺乳动物(gydF4y2BaegydF4y2Ba)。gydF4y2BafgydF4y2Ba,gydF4y2BaggydF4y2BaNPP的比例,直接被鸟类(gydF4y2BafgydF4y2Ba)和哺乳动物(gydF4y2BaggydF4y2Ba)。在gydF4y2BabgydF4y2Ba- - - - - -gydF4y2BaegydF4y2Ba、体重和能量估计个别鸟类和哺乳动物,与酒吧之和。误差表示95%置信区间来源于10000蒙特卡罗模拟法估计将身体质量的不确定性,人口密度,日常能量消耗方程,同化效率不同的食物类型,每个物种和NPP的饮食成分。在gydF4y2BafgydF4y2Ba,gydF4y2BaggydF4y2Ba,灰色酒吧表明NPP的直接消费,白色线条表示的百分比NPP间接支持鸟类和哺乳动物进食时消耗的平均营养级无脊椎动物被假定为2.5,其中的误差表示假定平均营养级的2.4和2.6。注意日志的规模gydF4y2BaygydF4y2Ba轴gydF4y2BafgydF4y2Ba,gydF4y2BaggydF4y2Ba。编号为gydF4y2Bad、egydF4y2Ba提供补充数据表1、2。gydF4y2Ba
通过消耗的总能量流是放大了所有精力充沛的通路的2.5倍(2.2 - -3.0;所有范围报告95%置信区间)相对于原始森林被砍伐的森林。在所有三种生境类型,总能量摄入量减少鸟类远远大于由哺乳动物(无花果。gydF4y2Ba2 d, egydF4y2Ba和扩展数据表gydF4y2Ba1gydF4y2Ba)。鸟类占67%、68%和90%的鸟类和哺乳动物的直接消费总量在原始森林,砍伐森林和油棕,分别。尽管哺乳动物生物量高于鸟生物量在古老和砍伐森林,新陈代谢每单位质量更高的鸟类,因为他们的小体型;因此,在能量和消费利率条款,鸟社区主导。仅靠鸟类总能量摄入增加2.6倍(2.1 - -3.2)相对于原始森林被砍伐的森林。这主要是由于增加了2.5倍(1.7 - -2.8)告诉读者食人植物foliage-gleaning(占主导地位的充满活力的途径),和大多数其他喂养公会也表现出更大的增加(无花果。gydF4y2Ba二维gydF4y2Ba和gydF4y2Ba3gydF4y2Ba)。然而,鸟总能量摄入的油棕滴回到原始森林中类似的水平,与多个公会的崩溃。对于哺乳动物,都有一个类似的2.4倍(1.9 - -3.2)增加在总消费从原始记录森林,但这在油棕种植园大幅下降。最引人注目的是5.7倍(3.2 - -10.2)陆地哺乳动物食草动物的重要性增加记录相对于原始森林。四个人原始森林网站显示鸟类和哺乳动物的能量始终低于记录森林(扩展数据图。gydF4y2Ba5gydF4y2Ba)。gydF4y2Ba

圆圈的大小表示能量流的大小,和颜色表明鸟类或哺乳动物。gydF4y2Ba年代gydF4y2Ba的物种数量;gydF4y2BaEgydF4y2BaESWI物种的冗余和索引,因此,韧性(比较高的值表明高冗余;见正文)。为了清楚起见,公会小能量流动没有显示,但在补充数据列出gydF4y2Ba4gydF4y2Ba。图片由j·本特利。gydF4y2Ba
NPP的一部分流经鸟和哺乳动物社区增加了2.1倍(1.5 - -3.0)相对于原始森林被砍伐的森林。很少有NPP增加记录相对于原始森林(无花果。gydF4y2Ba2gydF4y2Ba)因为NPP相对完整记录森林斑块的增加抵消了非常低的生产率更多结构退化前的日志平台等领域gydF4y2Ba14gydF4y2Ba,gydF4y2Ba15gydF4y2Ba。在油棕种植园,油棕水果占NPP的一大部分,虽然大部分的这些收获,从生态系统中删除gydF4y2Ba17gydF4y2Ba。NPP的比例,1.62%(-2.13% - 1.35)是直接在原始森林被鸟类和哺乳动物;(2.57 -5.07%),这一比例已升至3.36%记录森林但降到0.89%(0.57 -1.44%)棕榈油(无花果。gydF4y2Ba2 f, ggydF4y2Ba和扩展数据表gydF4y2Ba2gydF4y2Ba)。gydF4y2Ba
如果所有无脊椎动物吃食草动物或腐食者(也就是说,在一个营养级的2.0),和营养效率是10% (ref。gydF4y2Ba10gydF4y2Ba),NPP的总量支持鸟类和哺乳动物食物摄入量的组合将是9%,16%和5%的原始森林,砍伐森林和油棕,分别。然而,如果消耗的平均营养级无脊椎动物是2.5(即食草动物和食肉动物),相应的比例将为27%、51%和17%(图gydF4y2Ba2 f, ggydF4y2Ba)。告诉读者食人植物作为禽流感社区是占主导地位的喂养模式,这些数字是由鸟的饮食。鸟类的原始森林,0.35%的NPP支持直接食草性和frugivory,但是大约22%的NPP(假定无脊椎动物营养级2.5)是间接地告诉读者食人植物所需的支持。鸟类在森林砍伐的等效数字是0.83%和46%。因此,鸟类占更大的NPP的间接消耗。鸟的饮食研究在该地区的原始森林并记录表明,无脊椎动物消耗平均营养级的2.5 (ref。gydF4y2Ba18gydF4y2Ba;k·山姆,个人通信),这表明间接NPP消费的高端估计(在砍伐森林,50%左右)是合理的。gydF4y2Ba
比较有趣的是如此高的分数的NPP无脊椎食草性的直接估计。扫描这些森林的树落叶表明只有7.0%的树树冠叶面积(NPP)总量的1 - 3%是被树叶食草性gydF4y2Ba14gydF4y2Ba,gydF4y2Ba16gydF4y2Ba,但这样的估计不包括其他途径无脊椎动物,包括食草性的火情,地上和地下的刺,分层开采,水果,wood-feeding和树冠,垃圾和近地面层detritivory。无脊椎动物生物量的增加和食草性相比,森林砍伐原始森林曾被报道成雾的研究在这个格局gydF4y2Ba19gydF4y2Ba。如此之高的消费水平NPP的无脊椎动物可能会对生态系统植被生物量影响生产,建议,首先,无脊椎动物,食草性有一个实质性的影响从日志和恢复,其次,食虫鸟类密度可能施加实质性的间接控制生态系统的恢复。gydF4y2Ba
喂养公会之间的能量流动分布的生境类型(图中非常稳定。gydF4y2Ba3gydF4y2Ba),这表明砍伐森林的放大能量流动不扭曲整个脊椎动物群落的营养结构。整体鸟告诉读者食人植物饮食能量是由,引人注目的是不变的占66%,63%和66%的鸟类能量消耗在原始森林,砍伐森林和油棕,分别。Foliage-gleaning主导无脊椎动物消费的模式在所有三种类型的栖息地,大力frugivory是第二个最重要的喂养模式(分别为26%、27%和19%)。哺乳动物的饮食更均匀地分布在喂养公会,但frugivory(31%、30%、30%)和folivory(24%、38%、26%)占主导地位。小型哺乳动物的食虫动物可能是采样(见gydF4y2Ba方法gydF4y2Ba告诉读者食人植物)哺乳动物的贡献可能略大于估计。明显的相对大小恒常性的喂养通路完整和干扰的生态系统是值得注意的,而不是敏感生境类型之间的合理的喂养行为的变化(见gydF4y2Ba补充讨论gydF4y2Ba)。没有证据重大转变的主要喂养公会:校长喂养通路出现在森林的原始森林维护记录。gydF4y2Ba
当检查改变在物种水平在砍伐森林,鸟最大的绝对增加食品消费在树栖食虫类和杂食动物(无花果。gydF4y2Ba4gydF4y2Ba和扩展数据图。gydF4y2Ba2gydF4y2Ba)。特别是,这种变化表现为大诗人物种的丰度的增加(gydF4y2BaPycnonotusgydF4y2Baspp)。鸟类未显示出显著或大幅削减整体能源消耗。油棕种植园,鸟还不到总食品消费的砍伐森林,但类似于原始森林。然而,这是由非常高的少数物种的丰富,尤其是一个树栖杂食者(yellow-vented夜莺gydF4y2BaPycnonotus goiaviergydF4y2Ba)和三个树栖食虫类(gydF4y2BaMixornis bornensisgydF4y2Ba,gydF4y2BaRhipidura是gydF4y2Ba,gydF4y2BaCopsychus saularisgydF4y2Ba),而能量流经大多数其他鸟类都大大减少(图gydF4y2Ba4 bgydF4y2Ba和扩展数据图。gydF4y2Ba2 bgydF4y2Ba)。gydF4y2Ba

一个gydF4y2Ba,gydF4y2BabgydF4y2Ba能源消费变化相对于原始森林被砍伐的森林物种(gydF4y2Ba一个gydF4y2Ba)和油棕相对于原始森林(gydF4y2BabgydF4y2Ba)。20种经历最大的增加(红色)和减少(蓝色)在两种生境类型显示。鸟类所示较轻的语气和哺乳动物物种所示一个深色的基调。误差线表示95%的置信区间,源自10000年蒙特卡洛模拟估计将身体质量的不确定性,人口密度,日常能量消耗方程,同化效率不同的食物类型和组成的每一物种的饮食。gydF4y2Ba
对于哺乳动物来说,消费增加在砍伐森林是由消费主导的大型陆生食草动物增加了5.7倍(3.2 - -10.2),特别是黑鹿(gydF4y2Ba黑鹿单色的gydF4y2Ba)和亚洲象(gydF4y2BaElephas马克西姆斯gydF4y2Ba;无花果。gydF4y2Ba4gydF4y2Ba无花果和扩展数据。gydF4y2Ba2 bgydF4y2Ba和gydF4y2Ba3gydF4y2Ba),连同小杂食动物,主要是啮齿动物(原生带刺的老鼠,非黑老鼠;无花果。gydF4y2Ba4gydF4y2Ba)。一些雨林物种表现出强烈的下降(例如,大鼠鹿gydF4y2BaTragulus矮麝香鹿gydF4y2Ba和棕色带刺的老鼠gydF4y2BaMaxomys王侯gydF4y2Ba)。油棕,大多数哺乳动物崩溃(图。gydF4y2Ba4 bgydF4y2Ba)和有限的消费是由几个disturbance-tolerant栖息地多面手(例如,红色麂gydF4y2BaMuntiacus muntjakgydF4y2Ba,黑色的老鼠gydF4y2Ba家鼠gydF4y2Ba果子狸),尽管这些物种比原始森林中观察到的在较低密度(扩展数据图。gydF4y2Ba2gydF4y2Ba)。gydF4y2Ba
用很少的例外,放大能量流在被砍伐的森林似乎保留了相同级别的弹性如原始森林。物种的多样性和优势在任何途径可以衡量的韧性,通往物种的损失。我们评估各个通路通过定义一个精力充沛的精力充沛的主导地位Shannon-Wiener指数(ESWI)审查跨物种分布的能量流;低ESWI表示通路与高度依赖几个物种,因此潜在的脆弱性(图。gydF4y2Ba3gydF4y2Ba)。整个ESWI在公会不原始森林并记录之间的不同(gydF4y2BatgydF4y2Ba2,34gydF4y2Ba=−0.363,gydF4y2BaPgydF4y2Ba= 0.930),但从原始森林到棕榈油(大幅下降gydF4y2BatgydF4y2Ba2,34gydF4y2Ba=−3.826,gydF4y2BaPgydF4y2Ba= 0.0015),从记录森林油棕(gydF4y2BatgydF4y2Ba2,34gydF4y2Ba=−3.639,gydF4y2BaPgydF4y2Ba= 0.0025;线性mixed-effects模型、栖息地类型为固定效应和公会为随机效应;对模型系数看到补充表gydF4y2Ba3gydF4y2Ba)。gydF4y2Ba
因此,鸟类,物种的多样性导致主要精力充沛的途径维护从原始记录森林过渡但在油棕大幅下降。哺乳动物多样性和ESWI通常显示低于鸟类,但十喂养的公会维持或增加ESWI相对于原始森林被砍伐的森林,但油棕崩溃(无花果。gydF4y2Ba3gydF4y2Ba)。陆生食草性是最大的哺乳动物通路在被砍伐的森林,但依赖于只有四个物种和可能是最脆弱的大途径:几个大型哺乳动物(尤其是黑鹿)发挥主导陆生食草性作用的砍伐森林。同时,大胡子猪(gydF4y2BaSus barbatusgydF4y2Ba),唯一的野生suid在婆罗洲,形成的一个重要组件和功能独特的陆地杂食性通路。这些大型动物尤其敏感,人为压力比如打猎,或相关的致病压力就是明证最近急剧下降的大胡子猪在沙巴由于亚洲爆发猪瘟(数据收集)gydF4y2Ba20.gydF4y2Ba。gydF4y2Ba
脊椎动物种群在整个热带地区狩猎压力特别敏感gydF4y2Ba21gydF4y2Ba。我们的研究网站几乎没有狩猎,但灵敏度分析我们探索人口密度减少50%的精力充沛的后果这些物种可能受到影响的目标和/或不加选择地狩猎(扩展数据图。gydF4y2Ba4gydF4y2Ba)。有针对性的捕杀鸟类包括有商业价值的,和gun-hunted哺乳动物(胡子猪,有蹄类动物,野牛和哺乳动物的药用价值)。不加选择地狩猎物种包括鸟类和哺乳动物可能被困在网和陷阱。在砍伐森林狩猎降低鸟和哺乳动物能量流动,但仍让他们水平高于动物区系的完整的原始森林。这种狩猎带来鸟能量水平接近(但仍高于)的原始森林。为哺乳动物,然而,即使集中猎杀记录森林似乎比原始森林保持更高的能量流。因此,只有非常沉重的狩猎可能“抵消”放大能量在被砍伐的森林。gydF4y2Ba
我们砍伐森林的放大能量通路可能产生的自下而上的营养因素包括增加资源供给,适口性和可访问性。更开放的森林砍伐森林导致更多的植被结构接近地面gydF4y2Ba22gydF4y2Ba,gydF4y2Ba23gydF4y2Ba大通才食草动物,哺乳动物,因此更容易展示最显著增加哺乳动物的公会。竞争的优先级增加了植物的光,因此植被的快速增长策略在砍伐森林导致更高的叶片养分含量和降低叶化学防御食草性gydF4y2Ba24gydF4y2Ba,gydF4y2Ba25gydF4y2Ba以及更高的果期和开花率gydF4y2Ba19gydF4y2Ba和更大的聚集在资源供应gydF4y2Ba9gydF4y2Ba。这可能增加资源可用性和适口性支持高无脊椎动物和脊椎动物食草动物密度gydF4y2Ba25gydF4y2Ba。的行为干扰了生态系统从一个保守的化学防御状态更动态放大能量和营养流,但不是在某种程度上导致严重破坏动物群落组成。自上而下的营养因素也可能扮演一个角色在放大中间营养级的能量流动,通过等机制增加保护陆生或嵌套哺乳动物和鸟类从空中掠食者在茂密的植被地面层。这可能部分解释了大量的啮齿动物,但是几乎没有证据表明营养释放在这个网站,因为坚持高密度的哺乳动物食肉动物gydF4y2Ba26gydF4y2Ba。总的来说,更多的自底向上的机制和无脊椎动物消费激增表明增加资源供给和适口性很大程度上解释了放大的消费途径在砍伐森林。另一种可能性是,放大了脊椎动物的能量并不意味着放大整体动物能量,而是一个大的能量从无边无际的无脊椎动物捕食路径(例如,拟寄生物);这似乎不太可能,但需要进一步探索。gydF4y2Ba
油棕种植园显示一个大比例的下降NPP被哺乳动物和鸟类相比,砍伐森林gydF4y2Ba12gydF4y2Ba。哺乳动物种群崩溃,因为他们更脆弱,避免人类,也没有套哺乳动物多面手能够介入gydF4y2Ba27gydF4y2Ba,gydF4y2Ba28gydF4y2Ba。鸟类显示更温和下降,所观察到的类似的水平在原始森林,因为一套广泛的多面手物种能够适应和利用生境类型扰动梯度,而且因为体积小,流动呈现他们不太敏感的人类活动gydF4y2Ba29日gydF4y2Ba。有一致的油棕下降ESWI为哺乳动物,鸟类,尤其是指示大幅提高生态系统脆弱性在许多通路。gydF4y2Ba
总之,我们的分析展示了极大的研究记录的动态和生态活力的自然森林,甚至大量和反复登录在婆罗洲森林如发现。很可能的模式、机制和基本生态能量我们描述一般大多数热带森林;放大后的多个生态系统流程日志也被报道在肯尼亚砍伐森林gydF4y2Ba9gydF4y2Ba类似,但是需要详细分析一系列的热带森林阐明生物地理的的重要性,气候或其他因素。我们强调,我们的发现并不减少保护结构完整的原始森林的重要性,而是问题恶化的意思,一个新的生态价值的记录和其他结构退化的森林,加强他们的保护意义的议程gydF4y2Ba30.gydF4y2Ba。我们已经表明,物种的多样性不仅存在,而且在砍伐森林环境中茁壮成长。此外,这种生态活力可能提高生态系统结构性复苏的前景。的动物区系的完整无缺,我们研究景观接近最好的情况,因为狩猎压力较低。如果可以免受沉重defaunation砍伐森林,我们的分析表明,它们可以充满活力的生态系统,提供许多关键生态系统功能水平远高于原始森林。登录保护森林景观有着至关重要的作用在保护全球生物多样性和生态功能。gydF4y2Ba
方法gydF4y2Ba
领域的网站gydF4y2Ba
收集的数据记录森林在改变了森林生态系统的稳定性(安全)项目景观(4′43°N, 117°35′E)在沙巴,马来西亚gydF4y2Ba31日gydF4y2Ba低地镶嵌景观的砍伐森林和油棕种植园(无花果。gydF4y2Ba1gydF4y2Ba)。砍伐森林经历了一轮选择性伐木(删除113米gydF4y2Ba3gydF4y2Ba哈gydF4y2Ba−1gydF4y2Ba)在1970年代和额外累积体积37 - 66 mgydF4y2Ba3gydF4y2Ba哈gydF4y2Ba−1gydF4y2Ba在随后的旋转直到2000年代早期,这类似于意味着提取量152米gydF4y2Ba3gydF4y2Ba哈gydF4y2Ba−1gydF4y2Ba在一个更大的220000公顷面积在沙巴gydF4y2Ba32gydF4y2Ba,gydF4y2Ba33gydF4y2Ba。记录最严重情节已经登录四倍多,而不记录情节已经记录只有两次;因此,整个景观采伐强度高。油棕种植园的数据收集从毗邻的油棕房地产。数据为原始森林(无花果。gydF4y2Ba1gydF4y2Ba从Brantian-Tantulit原始森林保护区)收集(一个大片段的记录的森林景观),和其他三个原始森林保护区在沙巴:Maliau流域保护区(植被、鸟类和哺乳动物)的丹浓谷保护区(植被和鸟类)和Sepilok森林保护区(鸟类)。数据收集发生在2010年和2017年之间。张成的示例网站日志的梯度强度和生物量观测到整个景观(扩展数据图。gydF4y2Ba1gydF4y2Ba)。研究网站经历了非常低的狩猎的压力比其他地区的婆罗洲由于困难访问附近的城镇和文化因素,包括当地居民之间的相对有限的森林使用gydF4y2Ba27gydF4y2Ba。数据在每个物种调查或估计给出了补充数据gydF4y2Ba1gydF4y2Ba。gydF4y2Ba
植被NPP和调查gydF4y2Ba
NPP测量五个记录1公顷的土地安全项目区域的不同强度的日志(5年的数据),在四个原始森林Maliau盆地1公顷土地保护区(两个情节,4年的数据)和丹浓谷保护区(两块,2年的数据)gydF4y2Ba14gydF4y2Ba,gydF4y2Ba16gydF4y2Ba成熟,一个0.36公顷油棕情节(2年数据),在标准化的全球生态系统监测网络的协议gydF4y2Ba15gydF4y2Ba(无花果。gydF4y2Ba1gydF4y2Ba,扩展数据图。gydF4y2Ba1gydF4y2Ba和补充数据gydF4y2Ba3gydF4y2Ba)。我们量化以下NPP组件:伍迪NPP(茎,粗根和分支机构),树冠NPP(树叶、树枝和生殖器官)NPP和细根gydF4y2Ba10gydF4y2Ba。所有情节都至少有两个树茎和粗根NPP量化人口普查标准。树冠NPP(垃圾陷阱)和细根NPP(根长在肉内核心)监控月度和季度,分别。油棕种植园NPP估算是基于棕榈普查和异速生长高度,每月统计的花串,水果串连接和修剪的叶子结合一次性调查它们的质量,和季度收获根长在肉内的核心。gydF4y2Ba
哺乳动物的调查gydF4y2Ba
描述地面中型和大型non-volant哺乳动物社区,我们获得的检测/非探测数据从遥控数码相机陷阱(Reconyx HC500) 2011年5月到2017年12月gydF4y2Ba27gydF4y2Ba,gydF4y2Ba34gydF4y2Ba。相机陷阱被部署在882位置,分层在原始森林(gydF4y2BangydF4y2Ba= 236),被砍伐的森林(gydF4y2BangydF4y2Ba= 539)和油棕(gydF4y2BangydF4y2Ba= 107)。两个调查设计。第一个层次,集群设计,相机被23 - 232在网格(42877相机陷阱的夜晚,平均连续摄像机部署在49天gydF4y2Ba35gydF4y2Ba)。第二间隔与双摄像机系统的设计更广泛/景观车站> 1公里(11403相机陷阱的夜晚,相机平均连续部署在47天gydF4y2Ba34gydF4y2Ba)。在这两种情况下,相机被部署离地面20 - 50厘米,干扰植物保持到最低限度,没有鱼饵或诱惑。gydF4y2Ba
地面小型哺乳动物调查2011年5月到2014年7月使用国产钢丝网陷阱,部署在全球1488个地区分层在栖息地类型(432年、768年和288年在原始森林,砍伐森林和油棕,分别gydF4y2Ba35gydF4y2Ba)。陷阱的位置被聚集到1.75公顷捕获12×4位置的网格与23米间距。每个位置采样使用两个陷阱(间距为5 - 20 m)放置在或接近地面(0 - 1.5)和饵油棕果实。陷阱是连续七早上检查,捕捉到个人都使用皮下被动感应应答器标记在捕获位置之前被释放gydF4y2Ba27gydF4y2Ba,gydF4y2Ba35gydF4y2Ba。一些陷阱网格采样不止一次31网格(14),和总抽样工作是34058陷阱的夜晚。gydF4y2Ba
会飞的哺乳动物,我们蝙蝠从竖琴陷阱捕获数据集用于森林2011年4月至2012年6月gydF4y2Ba33gydF4y2Ba。蝙蝠被抓获在42个采样点的12个网站(3原始森林,砍伐森林9日),在陷阱设置50 - 150。7陷阱设置穿过森林小径和日志记录每晚打滑,然后第二天搬到一个新职位。蝙蝠是用独特的前臂乐队或翼活检之前释放这夺回个人可以从分析识别和移除。没有类似的数据可供油棕竖琴陷阱在开放的栖息地是无效的。gydF4y2Ba
鸟的调查gydF4y2Ba
禽流感点计数进行跨越森林和周围的356个地点油棕地产,与网站由180 - 220 (ref。gydF4y2Ba36gydF4y2Ba)。每个计算涉及到一个有经验的观察者(S.L.M.)记录所有物种所见所闻在下半场一段无限距离,包括鸟飞过。四项在每个站点之间凌晨和早上11点在2014年和2016年之间没有下雨。网站的平均时间间隔采样72天之间的第一个和最后一个访问。三个种类的迅速(gydF4y2BaAerodramus马克西姆斯gydF4y2Ba,gydF4y2Ba答:salanganagydF4y2Ba和gydF4y2Ba答:fuciphagusgydF4y2Ba),不能可靠地分离条件共同视为在大多数领域gydF4y2BaAerodramusgydF4y2Baspp。gydF4y2Ba
密度估计gydF4y2Ba
对于陆地中型和大型的哺乳动物,我们估计在每个相机陷阱密度点使用随机遭遇模型(REM)gydF4y2Ba37gydF4y2Ba38gydF4y2Ba。这种方法使用信息的大小相机陷阱检测区,和动物的运动速度,正确的捕获率数据(数量的动物通过单位时间)和密度估计。具体来说,REM包括所需的参数,为每一个物种:活动水平的比例(也就是说,24小时昼夜周期花活动和用于检测);活动时的移动速度;相机陷阱的有效探测角;相机陷阱的有效探测距离;和捕获率。活动水平的基础上估计的时间戳相机陷阱检测gydF4y2Ba39gydF4y2Ba和运动速度和检测区参数估计的基础上动物位置数据从相机陷阱的图像序列中恢复过来。这是可能的,因为我们的校准相机陷阱的位置(使用已知大小的一个对象,m杆)和特定的相机陷阱模型,我们使用(通过拍照的对象的大小在已知距离相机)。这允许我们恢复动物的距离和角度,从而估计图像序列中动物活跃时的速度gydF4y2Ba40gydF4y2Ba。有效探测角度和距离估计使用一个适应距离采样方法gydF4y2Ba39gydF4y2Ba。我们实现了快速眼动使用贝叶斯方法大型化,物种被视为随机效应和稀有物种,估计只有稀疏数据,成为可能的“借款”信息更常见的物种gydF4y2Ba41gydF4y2Ba。单独的大型化模型(包括土地利用类型作为协变量)活动的水平,速度和检测区参数被用来估计为每一个物种在每个土地利用后验分布。这些后验分布然后结合捕获率数据来估计密度、引导的数据提供估计的不确定性(补充数据gydF4y2Ba2gydF4y2Ba)。最后的密度估计广泛比得上预估数据在该地区的其他网站。gydF4y2Ba
估计陆地哺乳动物密度小gydF4y2Ba42gydF4y2Ba,我们使用空间显式再造型gydF4y2Ba43gydF4y2Ba。这个造型框架明确占一些个人与家庭范围的边缘捕获捕获网格可能并不总是可用的。空间显式再造型方法因此控制变异的有效采样面积捕获网格可能发生(例如,在扰动梯度)。单独的模型对于每个土地利用类型在R包secr安装gydF4y2Ba44gydF4y2Ba使用默认参数(即动物活动范围的泊松分布中心和half-normal检测函数),没有反是。缓冲区的陷阱周围100米的位置定义的模型集成。足够的数据来估计密度14种古老的小哺乳动物和砍伐森林。有太少的捕捉油棕允许模型拟合。gydF4y2Ba
与陆地哺乳动物,蝙蝠数据不通过重复测量设计。因此,密度的基础上派生一个20米检测半径(即0.126公顷)在每一个陷阱,和估计计算每个物种的总数量每累计检测区域每个栖息地类型。gydF4y2Ba
我们估计是当地丰富的鸟类的人均使用Royle-Nichols模型检测gydF4y2Ba45gydF4y2Ba。在分析之前,物种特异性检测的历史由池检测和非探测数据转换成离散的采样情况下根据现场参观。我们的建模框架描述丰富和检测使用分类habitat-specific拦截(原始森林,砍伐森林和油棕),将特有的斜坡和拦截,从常见的社区范围内分布随机效应。模型规范和检查程序后建立协议gydF4y2Ba34gydF4y2Ba。我们按比例缩小的模仿鸟类丰富(内个体数量的有效采样面积:7854米gydF4y2Ba2gydF4y2Ba每个点估算缓冲)密度每平方公里事后使用转换因子为0.785。gydF4y2Ba
专家判断和独立的估计gydF4y2Ba
对于一些其他哺乳动物不能可靠地采样通过相机陷阱或小动物陷阱来的例子,由于专性树栖栖息地的使用(一些灵长类动物和松鼠物种)或迁徙行为(例如,亚洲大象gydF4y2Ba大肠马克西姆斯gydF4y2Ba)——我们依靠估计基于遇到利率与这些物种实地考察过程中,或在独立研究在同一研究区域(补充数据gydF4y2Ba1gydF4y2Ba),例如,婆罗洲的猩猩(gydF4y2Ba彭哥pygmaeusgydF4y2Ba)gydF4y2Ba46gydF4y2Ba。亚洲象在砍伐森林和油棕密度估计的基础上观察到的行为的单一15-elephant群安全的景观,和原始森林的平均低密度在Maliau报告和其他沙巴原始森林的密度越高gydF4y2Ba47gydF4y2Ba。由于偏差引入的生活史高度移动的鸟类,模仿五种犀鸟的密度(gydF4y2BaAnthracoceros malayanusgydF4y2Ba,gydF4y2BaAnorrhinus galeritusgydF4y2Ba,gydF4y2BaBuceros犀牛gydF4y2Ba,gydF4y2BaRhinoplax守夜gydF4y2Ba,gydF4y2BaRhyticeros波形的gydF4y2Ba)以及伟大的argus (gydF4y2BaArgusianus argusgydF4y2Ba)和冠蛇雕(gydF4y2BaSpilornis cheelagydF4y2Ba从文献)纠正使用可用的信息。活动范围的估计每个犀鸟物种栖息地类型是围绕均值和扩展单位标准差。这是乘以465.3公顷的转换因子的基础上,意思是活动范围报告在七个物种(无线电遥测技术研究;补充数据gydF4y2Ba1gydF4y2Ba)计算扩展活动范围估计为每一个物种。每公顷密度估计被推断为扩大活动范围的倒数。这些大型鸟类总生态系统能量贡献了一个很小的部分,因此我们这些假设总体业绩非常不敏感。gydF4y2Ba
聚合栖息地类型gydF4y2Ba
当我们在类群结合数据,我们需要最大的抽样工作和社区的“最佳”的描述,我们聚合物种丰度估计每栖息地类型单一值。REM造型,数据对于一个给定的栖息地被用于模型来估计一个值所需的每个快速眼动参数(例如,速度、探测角度和距离,活动水平,捕获率和密度)在每个栖息地。因此,我们单位的复制是工会,没有空间组件在栖息地类型,因此没有空间自相关变量,可以探索。gydF4y2Ba
每天精力充沛的支出和食物摄取gydF4y2Ba
日常能量消耗为每个单独的物种从体重计算使用发表大型化异速生长的方程领域对哺乳动物和鸟类代谢率gydF4y2Ba48gydF4y2Ba(见补充表gydF4y2Ba1gydF4y2Ba方程和参数值)。每种食品饮食产生的分数分配给每个物种的基础上由三个合作者(O.R.W.专家专家的判断N.J.D. S.L.M.),和食物摄取率计算的基础上同化效率为每个喂养协会和食品类型gydF4y2Ba49gydF4y2Ba,gydF4y2Ba50gydF4y2Ba(补充表中总结gydF4y2Ba2gydF4y2Ba)。gydF4y2Ba
EWSIgydF4y2Ba
我们定义和使用ESWI检查能量流的分区跨物种通过类比Shannon-Wiener多样性指数是衡量种群丰度是如何跨物种传播gydF4y2Ba
在这gydF4y2BaegydF4y2Ba我gydF4y2Ba通过物种能量流的比例gydF4y2Ba我gydF4y2Ba在一个社区gydF4y2BangydF4y2Ba物种,通过社区相对于总能量流。gydF4y2Ba
比较在公会和栖息地类型gydF4y2Ba
测试在ESWI显著差异在栖息地类型,我们应用线性mixed-effects模型与ESWI作为响应变量,生境类型(原始森林,砍伐森林,油棕)作为固定效应和协会(gydF4y2BangydF4y2Ba= 18,看到补充数据gydF4y2Ba1gydF4y2Ba)作为随机效应(随机拦截),与方差结构允许不平等的方差在栖息地类型(varIdent函数,它实现了一个不同的方差为每个栖息地;模型结构方差比没有好,Akaike信息标准的120.5和128.3,分别),使用nlme包gydF4y2Ba51gydF4y2Ba。成对的事后比较栖息地,与图基的调整,进行了使用emmeans包gydF4y2Ba52gydF4y2Ba。gydF4y2Ba
不确定性的计算gydF4y2Ba
我们认为有以下的不确定性变量:体重的物种、人口密度、日常能量消耗(迪)方程,同化效率不同的食物类型,每个物种和NPP的饮食成分。身体质量,我们从一个截断正态分布(下限= 1 g)的意思是观察到的身体质量和标准偏差为15%。我们根据这个标准偏差为鸟类研究热带鸟类gydF4y2Ba53gydF4y2Ba和应用相同的哺乳动物的一致性为15%,没有其他数据gydF4y2Ba53gydF4y2Ba。人口密度,我们使用了10000引导人口密度的估计模型。此外,对于鸟类和蝙蝠(人口密度估计是基于探测半径在采样点),我们把半径的不确定性通过借鉴截断正态分布与标准偏差为20%,50%和150%的低和上界的估计半径。我们分配30%的不确定性的一些专家猜测的物种,有一个非常小的影响最后的结果。迪,我们估计的95%置信区间预测所述ref。gydF4y2Ba48gydF4y2Ba。同化效率,我们从一个随机的贝塔分布,使用平均值和标准偏差的食物类型和公会文学(补充表gydF4y2Ba2gydF4y2Ba)。部分饮食成分,我们生成了一个对称的贝塔分布,与20%的峰值不确定性时食品集团占50%的物种的饮食和不确定性时,食品集团占0%或100%的饮食。可能是日志记录和转换为油棕导致系统的变化对节肢动物的饮食成分。因此,我们还进行了两个额外的分析的部分消费节肢动物(在一个分析)和树叶(在其他分析)增加了30%的饮食好坏参半的物种。NPP的不确定性来自一个截断正态分布的平均值和标准偏差源自于字段数据gydF4y2Ba14gydF4y2Ba,gydF4y2Ba16gydF4y2Ba和上下边界的分布设置在均值±2个标准差。gydF4y2Ba
量化的不确定性估计NPP消耗的能量摄入和比例,我们跑10000模拟,取代原来的值计算值是从随机分布。首先,我们估计总同时假设所有组件的不确定性和不确定性的2.5%和97.5%百分位数值模拟计算获得95%置信区间估计。第二,量化每个变量导致了多少总不确定性,我们跑10000套模拟中只有一个变量的不确定性,而其他人则保持不变。我们计算的5%到95%百分位范围uncertainty-in-one-variable-at-the-time估计和uncertainty-in-all-variables估计,总不确定性和每个变量的贡献被认为是两者的比例gydF4y2Ba54gydF4y2Ba。gydF4y2Ba
绝对消费的不确定性估计由人口密度的不确定性和迪异速生长的主导消费者(扩展数据图。gydF4y2Ba6gydF4y2Ba)。不确定性在饮食比例分配,同化效率或消费异速生长的方程作出相对温和的贡献。因此,进一步减少不确定性是最好的有针对性的提高估计的丰度和生产更好的迪异速生长的几个主导消费者(扩展数据图。gydF4y2Ba7gydF4y2Ba),以及更好的评估等关键采样组小森林地面和树栖哺乳动物。计算NPP消耗的分数时,NPP估算的不确定性主导消费估计的不确定性,特别是在砍伐森林和棕榈油。gydF4y2Ba
警告gydF4y2Ba
在我们的分析有很多警告。一些类群可能是采样。这些包括几个小近地面层食虫动物(特别是鼩)不能可靠地检测到使用相机陷阱或fruit-baited住陷阱(陷阱陷阱需要漂移栅栏)和16个以果实为食的或nectivorous蝙蝠物种难以捕捉的热带森林火情gydF4y2Ba55gydF4y2Ba并有可能在某种程度上利用研究区。数据充分的树栖哺乳动物如灵长类动物和飞鼠估计从同一地区的其他研究(补充数据gydF4y2Ba1gydF4y2Ba)。我们没有测量NPP内原始森林的景观砍伐森林。然而,测量NPP的变化在原始网站在婆罗洲东北强烈不同土壤基质相当小(范围12.03 - -15.53毫克C公顷gydF4y2Ba−1gydF4y2Ba年gydF4y2Ba−1gydF4y2Ba;也就是说,±9%)gydF4y2Ba14gydF4y2Ba在原始森林,所以我们完全相信NPP安全景观是在这个范围内。我们的分析也不承担任何饮食变化在物种在扰动梯度(例如,无脊椎动物组成特定物种的相同分数的饮食是否在原始或砍伐森林),但灵敏度测试显示可能的饮食变化的影响可以忽略不计(gydF4y2Ba补充讨论gydF4y2Ba)。最后,我们假定一个动物的存在之间的相关性和消费的资源可能不会在所有情况下。如上所述,这些动物中观察到的碎片之间的油棕种植园可以通过记录和/或河岸的森林,因此我们估计在油棕的消费可能会高估。尽管如此,有少量的动物,如胡须的猪,猕猴和小型食肉动物有利于油棕补给区,大量的棕榈果和啮齿动物gydF4y2Ba27gydF4y2Ba,gydF4y2Ba56gydF4y2Ba。我们砍伐森林景观相邻是一个面积较大的适度砍伐森林北(无花果。gydF4y2Ba1gydF4y2Ba)。更加孤立和分散记录森林可能更defaunated比研究,即使没有狩猎,因此表现出较小的能量增加。gydF4y2Ba
报告总结gydF4y2Ba
进一步研究信息设计是可用的gydF4y2Ba自然投资组合报告总结gydF4y2Ba与这篇文章有关。gydF4y2Ba
数据可用性gydF4y2Ba
每个物种能量数据和快速眼动参数(哺乳动物)在补充数据是可用的gydF4y2Ba1gydF4y2Ba和gydF4y2Ba2gydF4y2Ba。gydF4y2Ba
代码的可用性gydF4y2Ba
的代码处理和统计分析的数据是可用的gydF4y2Ba补充的方法gydF4y2Ba。快速眼动分析代码可以从相应的作者合理请求或从裁判的补充方法。gydF4y2Ba38gydF4y2BagydF4y2Ba。gydF4y2Ba
引用gydF4y2Ba
吉布森,l . et al。主要森林对维持热带生物多样性是不可替代的。gydF4y2Ba自然gydF4y2Ba478年gydF4y2Ba,378 - 381 (2011)。gydF4y2Ba
Malhi Y。,Gardner, T. A., Goldsmith, G. R., Silman, M. R. & Zelazowski, P. Tropical forests in the Anthropocene.为基础。启包围。Resour。gydF4y2Ba39gydF4y2Ba,125 - 159 (2014)。gydF4y2Ba
Meijaard,大肠& Sheil d一片被砍伐的森林在婆罗洲,总比没有强。gydF4y2Ba自然gydF4y2Ba446年gydF4y2Ba974 (2007)。gydF4y2Ba
Bergius, M。,Benjaminsen, T. A., Maganga, F. & Buhaug, H. Green economy, degradation narratives, and land-use conflicts in Tanzania.世界开发。gydF4y2Ba129年gydF4y2Ba104850 (2020)。gydF4y2Ba
Giam X。,Clements, G. R., Aziz, S. A., Chong, K. Y. & Miettinen, J. Rethinking the ‘back to wilderness’ concept for Sundaland’s forests.医学杂志。Conserv。gydF4y2Ba144年gydF4y2Ba,3149 - 3152 (2011)。gydF4y2Ba
Burivalova, Z。,年代ekercioğlu, C. H. & Koh, L. P. Thresholds of logging intensity to maintain tropical forest biodiversity.咕咕叫。医学杂志。gydF4y2Ba24gydF4y2Ba,1893 - 1898 (2014)。gydF4y2Ba
语言,f M。,Frazão, F. S., Korasaki, V., Louzada, J. & Barlow, J. Identifying thresholds of logging intensity on dung beetle communities to improve the sustainable management of Amazonian tropical forests.医学杂志。Conserv。gydF4y2Ba216年gydF4y2Ba,115 - 122 (2017)。gydF4y2Ba
比克内尔,j·E。,Struebig, m . J。,Edwards, D. P. & Davies, Z. G. Improved timber harvest techniques maintain biodiversity in tropical forests.咕咕叫。医学杂志。gydF4y2Ba24gydF4y2BaR1119-R1120 (2014)。gydF4y2Ba
Schleuning, m . et al。森林碎片和选择性砍伐有不一致的影响在多个animal-mediated热带森林生态系统过程。gydF4y2Ba《公共科学图书馆•综合》gydF4y2Ba6gydF4y2Bae27785 (2011)。gydF4y2Ba
生态学的林德曼,r . l . trophic-dynamic方面。gydF4y2Ba生态gydF4y2Ba23gydF4y2Ba,399 - 417 (1942)。gydF4y2Ba
•奥德姆,h . t . &鸽子,r F。gydF4y2Ba热带雨林:研究辐照与生态El佛得角,波多黎各gydF4y2Ba。国家技术信息服务,维吉尼亚州斯普林菲尔德市(1970)。gydF4y2Ba
巴恩斯,公元et al。后果multitrophic热带土地利用的生物多样性和生态系统功能。gydF4y2BaCommun Nat。gydF4y2Ba5gydF4y2Ba5351 (2014)。gydF4y2Ba
里根,d p & Waide r B。gydF4y2Ba热带雨林的食物网gydF4y2Ba(芝加哥大学出版社,1996年)。gydF4y2Ba
Riutta, t . et al .日志扰动变化净初级生产力及其分配在婆罗洲的热带森林。gydF4y2Ba水珠。改变医学杂志。gydF4y2Ba24gydF4y2Ba,2913 - 2928 (2018)。gydF4y2Ba
Malhi, y . et al。全球生态系统监测网络:监测生态系统生产力和整个热带地区碳循环。gydF4y2Ba医学杂志。Conserv。gydF4y2Ba253年gydF4y2Ba108889 (2021)。gydF4y2Ba
Riutta, t . et al .重大和持续地下碳动力学的变化和在热带森林采伐后土壤呼吸。gydF4y2Ba水珠。改变医学杂志。gydF4y2Ba27gydF4y2Ba,2225 - 2240 (2021)。gydF4y2Ba
Kotowska, M . M。Leuschner C。Triadiati, T。,Meriem, S. & Hertel, D. Quantifying above- and belowground biomass carbon loss with forest conversion in tropical lowlands of Sumatra (Indonesia).水珠。改变医学杂志。gydF4y2Ba21gydF4y2Ba,3620 - 3634 (2015)。gydF4y2Ba
爱德华兹,d . p . et al .营养灵活性和持久性的林下叶层鸟类集中记录雨林。gydF4y2BaConserv。医学杂志。gydF4y2Ba27gydF4y2Ba,1079 - 1086 (2013)。gydF4y2Ba
削减大口水壶,r . m . et al .日志功能无脊椎动物在热带雨林的重要性。gydF4y2BaCommun Nat。gydF4y2Ba6gydF4y2Ba6836 (2015)。gydF4y2Ba
大口水壶,r . M。,Nathan, S. K. S. & Lee, P. A. K. African swine fever ravaging Borneo’s wild pigs.自然gydF4y2Ba593年gydF4y2Ba37 (2021)。gydF4y2Ba
Benitez-Lopez, a . et al .狩猎的影响在热带哺乳动物和鸟类种群。gydF4y2Ba科学gydF4y2Ba356年gydF4y2Ba,180 - 183 (2017)。gydF4y2Ba
Dobert, t·F。,Webber, B. L., Sugau, J. B., Dickinson, K. J. M. & Didham, R. K. Logging, exotic plant invasions, and native plant reassembly in a lowland tropical rain forest.《gydF4y2Ba50gydF4y2Ba,254 - 265 (2018)。gydF4y2Ba
Milodowski, d . t . et al .登录垂直树冠结构的影响在一个梯度的热带森林退化强度在婆罗洲。gydF4y2Baj:。生态。gydF4y2Bahttps://doi.org/10.1111/1365 - 2664.13895gydF4y2Ba(2021)。gydF4y2Ba
,s . et al .日志和土壤养分独立解释热带雨林植物性状表达。gydF4y2Ba新植醇。gydF4y2Ba221年gydF4y2Ba,1853 - 1865 (2019)。gydF4y2Ba
Swinfield, t . et al .成像光谱揭示了地形的影响和登录叶子化学热带森林的树冠的树。gydF4y2Ba水珠。改变医学杂志。gydF4y2Ba26gydF4y2Ba,989 - 1002 (2020)。gydF4y2Ba
布罗迪,j . f . &佐丹奴,a .缺乏营养释放大型哺乳动物的捕食者和猎物在婆罗洲。gydF4y2Ba医学杂志。Conserv。gydF4y2Ba163年gydF4y2Ba58 - 67 (2013)。gydF4y2Ba
Wearn, o . r . et al .哺乳动物物种丰度梯度的热带土地使用强度:层次结构大型化造型的方法。gydF4y2Ba医学杂志。Conserv。gydF4y2Ba212年gydF4y2Ba,162 - 171 (2017)。gydF4y2Ba
迪尔:j . et al .高碳股票森林为热带生物多样性提供有限的好处。gydF4y2Baj:。生态。gydF4y2Ba55gydF4y2Ba,997 - 1008 (2018)。gydF4y2Ba
米切尔,s . et al。森林砍伐的严重程度调节生物岛群岛的均化。gydF4y2Ba描述生态学gydF4y2Bahttps://doi.org/10.1111/ecog.05990gydF4y2Ba(2022)。gydF4y2Ba
雷诺兹,G。佩恩,J。Sinun, W。,Mosigil, G. & Walsh, R. P. D. Changes in forest land use and management in Sabah, Malaysian Borneo, 1990–2010, with a focus on the Danum Valley region.菲尔。反式。r . Soc。BgydF4y2Ba366年gydF4y2Ba,3168 - 3176 (2011)。gydF4y2Ba
大口水壶,r . m . et al .大规模的森林碎片实验:项目改变了森林生态系统的稳定性。gydF4y2Ba菲尔。反式。r . Soc。BgydF4y2Ba366年gydF4y2Ba,3292 - 3302 (2011)。gydF4y2Ba
费舍尔,B。,Edwards, D. P., Giam, X. & Wilcove, D. S. The high costs of conserving Southeast Asia’s lowland rainforests.前面。生态。环绕。gydF4y2Ba9gydF4y2Ba,329 - 334 (2011)。gydF4y2Ba
Struebig, m . j .等人gydF4y2Ba生态研究进展gydF4y2Ba48卷(eds伍德沃德,g·奥格尔曼& e . j .)Ch。3、183 - 224(学者,2013)。gydF4y2Ba
迪尔:j . et al。影响零毁林应承担的责任:森林质量和狩猎压力限制哺乳动物的持久性在支离破碎的热带风景。gydF4y2BaConserv。列托人。gydF4y2Ba13gydF4y2Bae12701 (2020)。gydF4y2Ba
Wearn, o . R。,Carbone, C., Rowcliffe, J. M., Bernard, H. & Ewers, R. M. Grain-dependent responses of mammalian diversity to land use and the implications for conservation set-aside.生态。达成。gydF4y2Ba26gydF4y2Ba,1409 - 1420 (2016)。gydF4y2Ba
米切尔,s . l . et al。河岸储备帮助保护森林鸟在油棕主导景观社区。gydF4y2Baj:。生态。gydF4y2Ba55gydF4y2Ba,2744 - 2755 (2018)。gydF4y2Ba
Rowcliffe, j . M。场,J。,Turvey, S. T. & Carbone, C. Estimating animal density using camera traps without the need for individual recognition.j:。生态。gydF4y2Ba45gydF4y2Ba,1228 - 1236 (2008)。gydF4y2Ba
Wearn, o . r . et al .估计动物物种密度社区使用获得的信息仅从相机陷阱。gydF4y2Ba生态方法。另一个星球。gydF4y2Ba13gydF4y2Ba,2248 - 2261 (2022)。gydF4y2Ba
Rowcliffe, j . M。凯斯,R。,Kranstauber, B., Carbone, C. & Jansen, P. A. Quantifying levels of animal activity using camera trap data.生态方法。另一个星球。gydF4y2Ba5gydF4y2Ba,1170 - 1179 (2014)。gydF4y2Ba
Rowcliffe, j . M。詹森,p。凯斯,R。,Kranstauber, B. & Carbone, C. Wildlife speed cameras: measuring animal travel speed and day range using camera traps.远程参议员生态。Conserv。gydF4y2Ba2gydF4y2Ba,84 - 94 (2016)。gydF4y2Ba
Kery, m &罗伊尔,j . A。gydF4y2Ba应用分层建模生态学gydF4y2Ba卷。1(爱思唯尔,2015)。gydF4y2Ba
查普曼,p . m . et al .年际动态和持久性的小型哺乳动物社区择伐在婆罗洲热带森林。gydF4y2BaBiodivers。Conserv。gydF4y2Ba27gydF4y2Ba,3155 - 3169 (2018)。gydF4y2Ba
Borchers, d . l . & Efford m . g .空间明确的最大似然方法再研究。gydF4y2Ba生物识别技术gydF4y2Ba64年gydF4y2Ba,377 - 385 (2008)。gydF4y2Ba
Efford m . secr:空间显式再模型。4.5.7 R包版本(2022)。gydF4y2Ba
罗伊尔,j . a . &尼科尔斯,j . d .估计从重复presence-absence丰富数据或点计数。gydF4y2Ba生态gydF4y2Ba84年gydF4y2Ba,777 - 790 (2003)。gydF4y2Ba
水手,d . j . i . et al .密度的婆罗洲的猩猩(gydF4y2Ba彭哥pygmaeus moriogydF4y2Ba严重退化的森林和油棕种植园在沙巴,婆罗洲。gydF4y2Ba点。j·阿特拉通。gydF4y2Ba81年gydF4y2Bae23030 (2019)。gydF4y2Ba
很,c & Yoganand k最近估计的亚洲象在婆罗洲揭示了一个更小的人口。gydF4y2BaWildl。医学杂志。gydF4y2Bahttps://doi.org/10.1002/wlb3.01024gydF4y2Ba(2022)。gydF4y2Ba
Nagy, k。,Girard, I. A. & Brown, T. K. Energetics of free-ranging mammals, reptiles, and birds.为基础。启减轻。gydF4y2Ba19gydF4y2Ba,247 - 277 (1999)。gydF4y2Ba
Bairlein, f的能量和养分利用效率在鸟类审查。在gydF4y2BaProc。22日国际鸟类国会gydF4y2Ba(eds亚当斯。gydF4y2Ba&gydF4y2BaSlotow, r·h·) 2221 - 2246国际鸟类联盟gydF4y2Bahttps://www.internationalornithology.org/PROCEEDINGS_Durban/Symposium/S37/S37.4.htmgydF4y2Ba(1999)。gydF4y2Ba
克罗克,D。哈特,。,Gurney, J. & McCoy, C.估计每日食物摄入量的野生鸟类和哺乳动物,gydF4y2BaDEFRA;gydF4y2Bahttps://www.hse.gov.uk/pesticides/resources/R/Research_PN0908.pdfgydF4y2Ba(2022)。gydF4y2Ba
ibsen Pinheiro, J。,B一个tes, D. & R Core Team. nlme: Linear and nonlinear mixed effects models. R package version 3.1–160 (2021).
披散下来,r . v .估计边际的含义,即最小二乘方法。1.7.3 R包版本gydF4y2Bahttps://CRAN.R-project.org/package=emmeansgydF4y2Ba(2022)。gydF4y2Ba
阅读,问:d . et al .热带鸟类有较少的变量的身体尺寸。gydF4y2Ba医学杂志。列托人。gydF4y2Ba14gydF4y2Ba,20170453 (2018)。gydF4y2Ba
Fatichi, et al。不确定性分区挑战气候变化的重要细节的可预测性。gydF4y2Ba地球的未来gydF4y2Ba4gydF4y2Ba,240 - 251 (2016)。gydF4y2Ba
Struebig, m . J。克里斯蒂,L。,P我o, D. & Meijaard, E. Bats of Borneo: diversity, distributions and representation in protected areas.Biodivers。Conserv。gydF4y2Ba19gydF4y2Ba,449 - 469 (2010)。gydF4y2Ba
爱,k . et al .大胡子猪(gydF4y2BaSus barbatusgydF4y2Ba)利用分散forest-oil棕榈景观在沙巴,马来西亚婆罗洲。gydF4y2BaWildl。Res。gydF4y2Ba44gydF4y2Ba,603 - 612 (2018)。gydF4y2Ba
Asner, g . p . et al .映射地上碳储量在马来西亚婆罗洲推进森林保护和恢复。gydF4y2Ba医学杂志。Conserv。gydF4y2Ba217年gydF4y2Ba,289 - 310 (2018)。gydF4y2Ba
Martin A . r .托马斯& s c碳含量在热带树木的重新评估。gydF4y2Ba《公共科学图书馆•综合》gydF4y2Ba6gydF4y2Bae23533 (2011)。gydF4y2Ba
确认gydF4y2Ba
本研究是安全项目的一部分,在经济上支持由英国自然环境研究理事会的人造热带森林项目(NE / K016407/1和NE / K016377/1),森达美基金会和蝙蝠保护国际。本文也是一个产品的全球生态系统监测网络(gem.tropicalforests.ox.ac.uk)提供培训和方法。Maliau盆地和丹浓谷管理委员会,皇家学会东南亚雨林研究伙伴关系,沙巴基金会Benta Wawasan,国务秘书,沙巴首席部长的部门,沙巴林业部,沙巴生物多样性委员会和经济规划部门承认的支持在沙巴和访问网站。我们感谢k .山姆和r·邓恩对部分分析有用的建议。Y.M.杰克逊基金和欧洲研究委员会高级研究员格兰特,GEM-TRAIT (321131)。时被一个利华休姆信托赞助支持研究领导奖,和R.M.E.无关基金会的支持。gydF4y2Ba
作者信息gydF4y2Ba
作者和联系gydF4y2Ba
贡献gydF4y2Ba
Y.M.构思的分析,论文的写作与输入顶替,O.R.W.,N.J.D.,Z.G.D.,R.M.E. and M.J.S. O.R.W. developed the REM modelling; O.R.W., N.J.D., S.L.M. and M.J.S. collected and analysed the animal data; T.R. collected and analysed the vegetation data and worked on energetic data analysis. R.M.E. conceived and implemented the SAFE Project. H.B., N.M. and R.N. assisted in collection of field data with supervision from Y.M., Z.G.D., R.M.E. and M.J.S. All authors commented on the draft.
相应的作者gydF4y2Ba
道德声明gydF4y2Ba
相互竞争的利益gydF4y2Ba
作者宣称没有利益冲突。gydF4y2Ba
同行评审gydF4y2Ba
同行审查的信息gydF4y2Ba
自然gydF4y2Ba谢谢菲利浦-弗兰卡,卡米尔Piponiot Pieter Zuidema,另一方面,匿名的,审稿人(s)为他们的贡献的同行评审工作。gydF4y2Ba同行审查报告gydF4y2Ba是可用的。gydF4y2Ba
额外的信息gydF4y2Ba
出版商的注意gydF4y2Ba施普林格自然保持中立在发表关于司法主权地图和所属机构。gydF4y2Ba
扩展数据数据和表gydF4y2Ba
扩展数据图1的取样位置分布的梯度测井强度研究中,特征使用地上部干重(吨/公顷)。gydF4y2Ba
我们估计从spatially-explicit生物质表面的碳密度(30 m分辨率)源自机载光探测和测距数据(见(激光雷达)gydF4y2Ba57gydF4y2Ba完整的抽样细节)和碳转化为干重使用转换因子为0.47 (gydF4y2Ba58gydF4y2Ba)。提供一个具有代表性的当地生境条件,生物质提取从100米半径平均值缓冲区在每个采样点。在这个决议有广泛的在老干重值增长和砍伐森林,但平均值明显区分。左上的柱状图显示了生物和栖息地类型提取在每个采样位置对所有焦点类群的总和。taxa-specific情节简单地将信息分解到每个焦点组:植被初级生产力的采样点(点左上的面板的顶部),蝙蝠(右上图),鸟类(左下)和陆地哺乳动物(右下角)跨梯度。gydF4y2Ba
扩展数据图2占主导地位的鸟类和哺乳动物的食物能量消耗在原始森林,砍伐森林和油棕种植园。gydF4y2Ba
了解鸟类资源消耗(a - c)和哺乳动物(d-f)(前20名消费者在每一个森林类型)。误差表示95%的置信区间,源自10000年蒙特卡洛模拟估计将身体质量的不确定性,人口密度,日常能量消耗方程,同化效率不同的食物类型和组成的每一物种的饮食。数据所有物种的研究提供补充数据1。gydF4y2Ba
扩展数据图3变异能量摄入的鸟类和哺乳动物的大小类。gydF4y2Ba
鸟类(a)和哺乳动物的直接能量摄入(b)由身体质量类(对数刻度)在古老的森林生长(OG),砍伐森林和油棕种植园(凤凰社)。酒吧旁边的数字表示每个类的物种数量。误差表示95%的置信区间,源自10000年蒙特卡洛模拟估计将身体质量的不确定性,人口密度,日常能量消耗方程,同化效率不同的食物类型和组成的每一物种的饮食。数据提供的补充数据gydF4y2Ba4gydF4y2Ba。gydF4y2Ba
扩展数据图4的模拟狩猎总生物量的影响和鸟类和哺乳动物的能量摄入。gydF4y2Ba
体重的鸟类哺乳动物(a)和(b)和充满活力的食物摄入量的鸟类(c)和哺乳动物(d)在古老的森林生长(OG,深灰色,没有狩猎)和砍伐森林里(浅灰色)四种不同的狩猎场景:观察低狩猎压力(基线)和模拟人口密度降低50%的目标猎物物种,肆意猎杀物种有针对性和肆意捕杀的物种。有针对性的捕杀鸟类包括有商业价值的,和gun-hunted哺乳动物(胡子猪,有蹄类动物,野牛和哺乳动物的药用价值)。不加选择地狩猎物种包括鸟类和哺乳动物可能被困在网和陷阱。为每个类别的物种列表看到补充数据gydF4y2Ba1gydF4y2Ba。注意,这不是一个详尽的分析研究区域,而是一个狩猎压力说明估计的潜在影响营养能量上狩猎。捕杀鸟类目标可能包括鸟类的13%,占17%的鸟类体重和14%的鸟类观察下能耗低狩猎的压力。有针对性的捕杀哺乳动物可能包括10%的哺乳动物物种,占体重的46%,42%的哺乳动物观察下能耗低狩猎的压力。肆意捕杀鸟类可能包括鸟类的72%,占78%的鸟类体重和82%的鸟类观察下能耗低狩猎的压力。肆意捕杀哺乳动物可能包括哺乳动物物种的22%,占2%的哺乳动物体重和哺乳动物能源消耗的2%。狩猎压力同时应用,捕杀鸟类可能包括物种的86%,95%的鸟类体重和96%的鸟类观察下能耗低狩猎的压力,和捕杀哺乳动物可能包括32%的哺乳动物,哺乳动物体重48%的哺乳动物和44%的能源消耗压力下观察到低打猎。数据提供的补充数据gydF4y2Ba6gydF4y2Ba。gydF4y2Ba
扩展数据图5鸟类和哺乳动物生物量的变化和能量摄入个人原始森林站点和日志记录强度。gydF4y2Ba
体重和鸟类物种丰富度(a)和哺乳动物(b)和充满活力的食物摄入量的鸟类(c)和哺乳动物(d)在古老的森林(OG)增长,砍伐森林和油棕种植园(凤凰社)。OG森林数据分析分别由四个噩网站为哺乳动物鸟类和两个站点(见图gydF4y2Ba1gydF4y2Ba森林地图),记录的数据被分成两次登录,登录区域。对于哺乳动物来说,只有物种研究使用相机陷阱和竖琴陷阱包括(63%、63%和77%的哺乳动物物种和53%,45%和63%的总能量食物摄入量在噩,砍伐森林和OP,分别)。误差是95%置信区间源自10000年蒙特卡洛模拟估计将身体质量的不确定性,人口密度,日常能量消耗方程,同化效率不同的食物类型和组成的每一物种的饮食。数据提供的补充数据gydF4y2Ba7gydF4y2Ba。gydF4y2Ba
扩展数据图6能量摄入量的估计的不确定性的来源。gydF4y2Ba
能量摄入的不确定性来源的贡献比例(a)和净初级生产力(NPP)消耗(b)的鸟类和哺乳动物的栖息地类型的旧增长森林(OG),砍伐森林和油棕种植园(凤凰社)。我们认为有以下的不确定性变量:体重的物种、人口密度、日常能量消耗(迪)方程,同化效率不同的食物类型,每个物种的饮食,馏分组成和NPP。不确定性估计来自10000年蒙特卡洛模拟,和每个变量的贡献通过运行仿真评估总不确定性假设所有变量的不确定性同时在一个变量。数据提供的补充数据gydF4y2Ba5gydF4y2Ba。gydF4y2Ba
扩展数据图7的比例uncertainity总能量吸收由个别鸟类或哺乳动物物种。gydF4y2Ba
总不确定性由每个物种的比例排名能耗的鸟类(a)和哺乳动物(b)。gydF4y2Ba
补充信息gydF4y2Ba
补充信息gydF4y2Ba
这个文件包含补充讨论,引用,表1 - 3和描述的补充数据。gydF4y2Ba
补充的方法gydF4y2Ba
详细评论R代码的分析。代码中使用的所有数据都可以在补充数据文件。gydF4y2Ba
权利和权限gydF4y2Ba
开放获取gydF4y2Ba本文是基于知识共享署名4.0国际许可,允许使用、共享、适应、分布和繁殖在任何媒介或格式,只要你给予适当的信贷原始作者(年代)和来源,提供一个链接到创作共用许可证,并指出如果变化。本文中的图片或其他第三方材料都包含在本文的创作共用许可证,除非另有说明在一个信用额度的材料。如果材料不包括在本文的创作共用许可证和用途是不允许按法定规定或超过允许的使用,您将需要获得直接从版权所有者的许可。查看本许可证的副本,访问gydF4y2Bahttp://creativecommons.org/licenses/by/4.0/gydF4y2Ba。gydF4y2Ba
关于这篇文章gydF4y2Ba

引用这篇文章gydF4y2Ba
Malhi Y。,Riutta, T., Wearn, O.R.et al。gydF4y2Ba砍伐热带雨林有放大和多样的生态系统能量。gydF4y2Ba自然gydF4y2Ba612年gydF4y2Ba,707 - 713 (2022)。https://doi.org/10.1038/s41586 - 022 - 05523 - 1gydF4y2Ba
收到了gydF4y2Ba:gydF4y2Ba
接受gydF4y2Ba:gydF4y2Ba
发表gydF4y2Ba:gydF4y2Ba
发行日期gydF4y2Ba:gydF4y2Ba
DOIgydF4y2Ba:gydF4y2Bahttps://doi.org/10.1038/s41586 - 022 - 05523 - 1gydF4y2Ba
评论gydF4y2Ba
通过提交评论你同意遵守我们的gydF4y2Ba条款gydF4y2Ba和gydF4y2Ba社区指导原则gydF4y2Ba。如果你发现一些滥用或不符合我们的条件或准则请国旗是不合适的。gydF4y2Ba