








摘要
现代人类已经在欧洲居住了45000多年1,2.然而,我们对古代狩猎采集者的遗传相关性和结构的了解是有限的,因为那个时期的人类遗骸很少,分子保存也很差3..在这里,我们分析了356个古代狩猎采集者基因组,包括来自欧亚大陆西部和中部14个国家的116个人的新基因组数据,时间跨度在3.5万至5000年前。我们在来自西欧的旧石器时代晚期的格拉韦特人群体中鉴定出了一个遗传祖先剖面,这与中欧和南欧与这一考古文化相关的同时代群体截然不同4,但类似于与Aurignacian文化相关的先前个体。这一祖先特征在末次盛冰期(2.5万至1.9万年前)在欧洲西南部与梭鲁特文化有关的人类种群中幸存下来,并在末次盛冰期后重新向东北扩张的马格达伦文化中幸存下来。相反,我们揭示了南欧的基因更替,这表明在末次盛冰期前后,人类群体在当地发生了置换,伴随着与Epigravettian文化相关的人口从北到南的分散。至少从14000年前开始,与这种文化相关的祖先从南方传播到欧洲其他地区,在很大程度上取代了与抹大拉人相关的基因库。在跨越中石器时代开始的一段有限的混合时期之后,我们发现西欧和东欧狩猎采集者之间的遗传相互作用,他们也以表型相关变异的显著差异为特征。
主要
现代人至少在6万年前(ka)离开了撒哈拉以南的非洲,在他们最初向欧亚大陆扩张的过程中,他们在基因上与尼安德特人混合,导致今天大多数非非洲人口中有2-3%的尼安德特人血统5.基因组数据显示,现代人类出现在欧亚大陆西部1,2至少45 ka。来自超过40 ka的早期群体中的一些进一步与尼安德特人混合,正如来自保加利亚的Bacho Kiro(与最初的旧石器时代晚期(IUP)考古文化有关)和罗马尼亚的pettera cu Oase的个体最近渗透的信号所显示的那样2,6.来自那个时期的其他个体,如来自捷克的Zlatý k实验者和来自俄罗斯的Ust’ishim,并没有比其他非非洲群体携带更多的尼安德特人血统1,7这表明了尼安德特人与早期现代人在欧亚大陆最初扩张时的不同互动。然而,令人惊讶的是,这些40 ka以前的个体没有一个在今天欧亚人口的基因构成中留下实质性的痕迹1,2,6,7.携带祖先的最古老的基因组主要来自导致今天欧洲人的谱系是Kostenki 14(来自37 ka,与俄罗斯西部的考古关联不确定),Goyet Q116-1 (35 ka,与比利时的aurignacian相关)和Bacho Kiro 1653 (35 ka,可能来自保加利亚的aurignacian相关)2,4,8.这些数据表明,迄今为止分析的40 ka以前的个体的遗传祖先大部分已经灭绝或被随后的扩张同化1,9.Kostenki遗传标记(与Kostenki 14基因组相关,下文称为Kostenki群集或祖先)促成了后来的vstonice遗传群集(下文称为vstonice群集或祖先),以捷克Dolní vstonice遗址命名4.这种基因特征在中欧和南欧考古定义的格拉韦特文化(33-26 ka)的个体中是共同的,似乎在末次极盛期之后消失了4高强)。然而,来自西欧的同时期gravettian相关个体的遗传特征仍然未知,他们对LGM之后的种群的贡献也是未知的。LGM被认为是上一个冰河时代最冷的阶段,被认为是造成欧洲大部分地区人口下降的原因10例如,伊比利亚半岛和法国南部的梭鲁特文化(24-19 ka)就证明了这一点。在这一时期,其他被提议的人类生存的气候避难所是意大利半岛、巴尔干半岛和欧洲东南部平原,但这些地区的人口对后lgm欧洲人的实际遗传贡献存在很大争议11,12,13.
LGM之后,一个与来自比利时的Goyet Q116-1个体有遥远联系的遗传成分,可追溯到35 ka -命名为GoyetQ2祖先(以下,GoyetQ2群或祖先)-重新出现在与Magdalenian文化相关的西南和中欧的个体中(19-14 ka从伊比利亚到东欧,穿越中欧),并以混合形式出现在随后的最终旧石器时代和中石器时代的狩猎采集者中4,14但这一祖先的地理分布仍不清楚。相反,在南欧,早在17ka时,就在与Epigravettian文化相关的个体中发现了独特的狩猎-采集遗传图谱15(24-12 ka,从意大利半岛到横跨巴尔干半岛的欧洲东南部平原)。这个“Villabruna”祖先(以后,Villabruna集群或祖先)显示出与古代和现代近东人口的联系4,16但它向意大利半岛扩张的模式和速度仍未被探索。Villabruna祖先后来出现在中欧,被认为在很大程度上取代了与GoyetQ2祖先有关的群体4.然而,它的形成、传播和与来自东欧的同时代狩猎采集者的相互作用,以及他们与后来来自东南欧的新石器时代农民的相互作用,并没有很好地描述。
在这项研究中,我们分析了356个古代狩猎-采集者基因组,包括116个可追溯到35-5 ka的新基因组数据,以及一种基于纯合子度的新污染估计方法。我们系统地描述了从旧石器时代早期开始横跨欧亚大陆西部和中部的狩猎采集群体所经历的基因组转变,以及这些转变如何与文化和气候变化联系在一起。
古代DNA数据生成
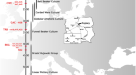
我们生成了102个新报告的狩猎采集者的全基因组测序数据,并增加了14个先前发表的个体的覆盖范围4.这些数据涵盖了大约3万年的时间跨度,从旧石器时代晚期到新石器时代晚期(这里的定义是陶器的存在,而不是农业自给经济,如果没有注明的话),来自多个史前文化背景,来自14个国家的54个考古遗址:1个来自比利时的奥里格纳古血统个体和1个来自罗马尼亚(35-33 ka)的文化未归属个体,15个来自西班牙、法国、比利时、捷克和意大利(31-26 ka)的gravetian血统个体,2个来自西班牙和法国(23-21 ka)的梭鲁特血统个体,9个来自法国、德国和波兰(18-15 ka)的magdalenian血统个体,4个来自意大利(17-13 ka)的epigravetian血统个体,2个来自德国(14 ka)的federmesser血统个体,81名来自欧亚大陆西部的中石器时代到新石器时代的觅食者(11-5 ka),以及1名来自塔吉克斯坦的欧亚大陆中部新石器时代的个体(8 ka)。1,扩展数据表1、补充数据1.一个,补充信息,章节1和补充图。1).
我们为每个个体建立了1到8个单链和双链基因文库,并在124万个单核苷酸多态性上对人类DNA进行了富集6(SNPs),然后对其进行测序,结果显示目标SNPs的平均覆盖率为0.04- 7.64倍。基因性别鉴定结果显示,男性78只,女性38只。12).现代人类DNA的污染水平是在线粒体DNA (mtDNA)、X染色体和常染色体DNA的基础上估计的,并使用单倍型复制模型,在纯合子(ROH)运行中扩展到常染色体数据(方法,补充信息,章节)2而且3.,补充图。2- - - - - -11及补充表1).对被严重污染的文库和被轻微污染的单独分析基因组文库进行过滤,以保持显示死后DNA损伤的读数(方法和补充图。10而且11).通过在每个位置随机采样一个等位基因,对目标snp进行伪单倍体基因型调用,结果在124万snp面板上覆盖了6,600 - 107万个snp(扩展数据表)1及补充资料1.一个).新生成的基因型与240个已发表的古代狩猎采集者基因组和现代世界范围内的人群合并,用于下游分析(补充数据)1.G).与Fu等人的建议相反。4但与Petr等人的观点一致。17,我们没有观察到随着时间的推移,大多数欧洲狩猎采集者的尼安德特人血统大幅减少(补充信息,章节6和补充图。15- - - - - -17).这进一步支持了一个模型,即现代人类的尼安德特人血统在他们渗入后没有长期下降18.
LGM之前
Gravettian文化是LGM之前在欧亚大陆西部分布最广泛的旧石器时代晚期文化之一19.它通常被认为是一种泛欧洲的文化马赛克,从材料到象征性作品都有区域差异20.,21.在这个有争议的框架下,根据颅骨测量和基因组数据,gravettian相关个体被认为代表了一个生物上同质的群体4,22.然而,已发表的gravettian相关基因组来自中欧和南欧,这使得来自西欧和西南欧洲的gravettitian相关人类群体的遗传特征未被描述。
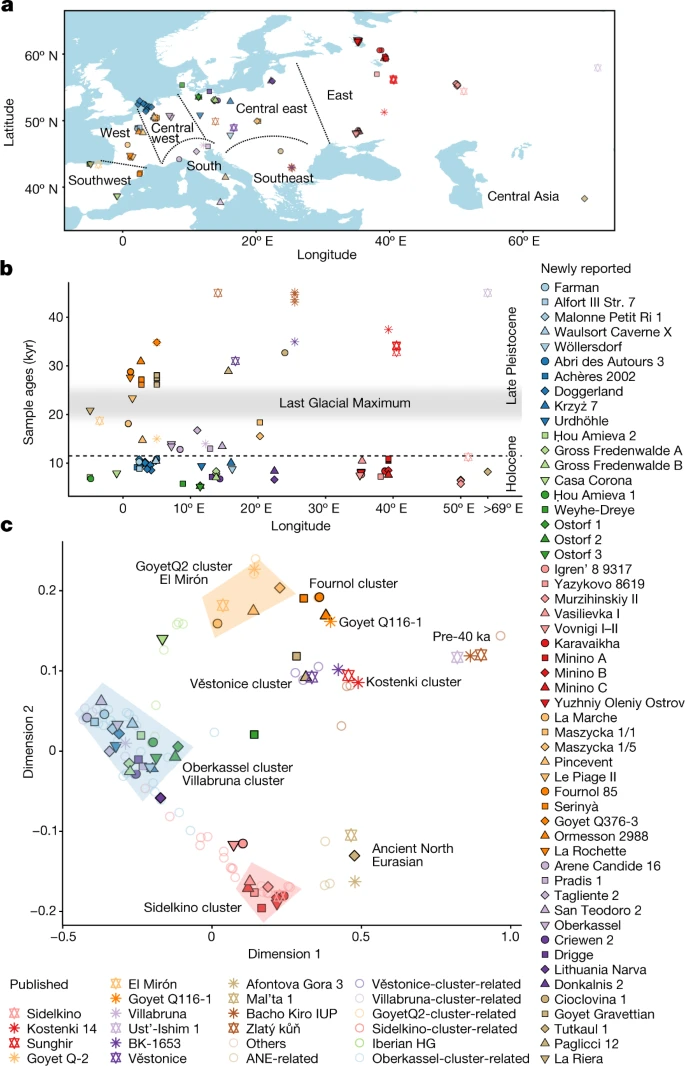
为了获得LGM之前欧洲狩猎采集者的基因组背景的概述,我们使用多维标度(MDS)绘制了成对外群的不相似矩阵f3.- 1−的统计信息f3.(穆布;pop1, pop2)2).这幅图揭示了三个不同群体的存在:(1)来自Ust’ishim、Bacho Kiro、Zlatý kolo冷藏和pettera cu Oase遗址的40 ka前群体,(2)来自中欧和南欧遗址的vstonice群,包括与gravettian相关的个体(Dolní vstonice、Pavlov、Krems-Wachtberg、Paglicci和Ostuni),以及(3)来自西欧和西南欧洲遗址(Ormesson、La Rochette、Fournol和两个Serinyà洞穴遗址(Mollet III和Reclau Viver))。先前描述的vstonice群集,包括新近报道的来自意大利南部Paglicci洞穴(Paglicci 12)的2.9万年前的个体,与先前发表的来自俄罗斯西部Sunghir和Kostenki 12的基因组密切相关,分别可追溯到34 ka和32 ka4,23.新定义的Fournol集群与来自比利时的aurignacian相关个体密切相关,其历史可追溯至35 ka (Goyet Q116-1和新报道的Goyet Q376-3个体)。值得注意的是,与Fu等人的报告相反。4,来自中欧的另一个与gravetian相关的人群(比利时的Goyet,n= 6个个体)在地理上和遗传上都介于vstonice和Fournol群之间。Goyet Q116-1和Goyet Q376-3与Fournol聚类之间的相似性也在mtDNA水平上被观察到,两组都包括携带mtDNA单倍群M的个体,这在LGM后的欧洲个体中没有发现24(扩展数据图。1而且2).
一个, lgm前个体MDS图。40 ka以前的群以及Fournol和vstonice群被标记为不同颜色的阴影区域。个体和群体用图中相同的颜色和符号绘制。1符号旁边有名字。b与gravettian相关的个体形成了两个不同的群体,中欧和南欧的个体作为vstonice集群的一部分,西欧和西南欧洲的个体作为Fournol集群的一部分。在中欧,来自Goyet的gravettian相关个体对这两个集群都表现出亲和力。误差条显示为1× s.e.m。(黑色)或3× s.e.m。(灰色)的f45厘米块叠刀分析估计的值。c,使用qpGraph创建的主要前lgm欧洲狩猎-采集血统的混合图建模。
我们进一步验证了在MDS图中观察到的vstonice簇和Fournol簇之间的遗传差异f4-statistics(补充数据2.B).所有属于Fournol类群的个体对Goyet Q116-1的亲和度均高于Sunghir类群(n= 4), vstone -cluster个体与Sunghir类群的亲缘性高于Goyet Q116-1(扩展数据图。3.).这些f4统计数据还证实,Goyet Q376-3与Goyet Q116-1具有相似的祖先,Kostenki 12与Sunghir类群具有相似的祖先,而来自保加利亚的Bacho Kiro 1653 (35 ka),来自罗马尼亚的Muierii 1 (34 ka)和Cioclovina 1 (32 ka),以及来自意大利南部的Paglicci 133 (33 ka)与Goyet Q116-1和Sunghir类群具有相同的亲缘关系。我们进一步测试了vstonice和Fournol聚类中的个体是否与这两个聚类的主要代表具有相似的等位基因频率。通过统计数据f4(Mbuti, Fournol 85;vstonice, test)和f4(穆,Věstonice;Fournol 85, test),我们表明所有vstone -cluster个体显著接近(|Z|>3)到vstonice组(n= 5)和Fournol-聚类个体更接近Fournol 85,而地理上处于中间位置的gravettian -相关Goyet组对这两个聚类都表现出额外的亲缘关系(图5)。2 b).
我们进一步用qpGraph对lgm前个体的遗传谱进行了建模(补充信息,章节)10和补充图。19- - - - - -25).外加剂图显示Bacho Kiro IUP组(n= 3)与多个早期现代人类谱系共享祖先2(补充资料,章节7),以及超过4.5万年前的Zlatý kzhevian基因组1是迄今为止最具差异的非非洲血统序列(扩展数据图。4).这也由f4-表单统计信息f4(姆布蒂,Zlatý ksergen;test1, test2),这与所有其他lgm前的狩猎采集者的零一致(补充数据2.摄氏度),表明Zlatý k实验者与被测群体的等距离关系。当gravettian相关的个体也包括在Kostenki 14的混合图中,我们发现Fournol 85最适合作为Goyet Q116-1的姐妹谱系,而vstonice类群则被模型为与Sunghir类群相关的谱系和与Goyet Q116-1 - Fournol 85分支相关的谱系的混合(图2)。2摄氏度).这也是由f4-表单统计信息f4(Mbuti, Fournol 85;Sunghir,测试),这对于vstonice集群中包括的所有个体都是显著阳性的(补充数据2.B).因此,如前所述2在美国,v斯通星系团本身是西方和东方血统的混合,这可能有助于在gravettian相关个体中观察到的颅骨形态的同质性22.
这些结果表明,在迄今为止所研究的格拉韦提相关人群中,存在于欧洲约40 - 30 ka之间的一些基因组祖先存活了下来,但不是全部。Kostenki(和Sunghir组)祖先为之前描述的v斯通群做出了贡献,该群由来自中欧和南欧的gravettian相关个体代表4.相比之下,Goyet Q116-1基因图谱在西欧和西南欧洲的gravettian相关个体中发现了新描述的Fournol集群。值得注意的是,这种遗传差异与来自欧洲不同地区的遗传分析的gravettian相关个体的太平间实践的差异相吻合。欧洲西部和西南部与Fournol群相关的个体一直沉积在洞穴遗址中,偶尔会出现人为痕迹,而与vstonice群相关的个体则分别与墓葬和/或个人饰品和赭石一起埋葬在中欧和南欧的露天或洞穴遗址中(补充图)。29- - - - - -32及补充表4).Fournol星团中最古老的个体是来自法国东北部的Ormesson 2988 (31 ka, Gravettian早期/中期),而来自比利时Goyet的Gravettian星团(27 ka, Gravettian晚期)被发现是vstonice和Fournol星团的混合体。这表明,在早期/中期和晚期格拉维特时代之间,与v斯通有关的祖先从东向西扩张,到达了中欧-西欧,并在这两个基因不同的前lgm人群之间形成了纵向混合斜群。
欧洲西南部和西部的LGM
梭鲁特文化在时间上介于Gravettian文化和Magdalenian文化(或Badegoulian文化)之间,发现于欧洲西南部和西部,被认为是LGM时期人类人口的气候避难所25,26.然而,与梭鲁特文化相关的群体在多大程度上与来自同一地区的早期和晚期人群具有遗传连续性尚不清楚,因为以前没有报道过来自梭鲁特文化相关个体的基因组数据。来自梭鲁特人相关个体(来自法国西南部的Le Piage II (23 ka)和来自西班牙北部的La Riera (14, 21 ka))的新测序基因组都显示出与Fournol和GoyetQ2外群成员的普遍亲缘关系f3.-statistics(补充数据2.一个).在MDS图中,Le Piage II个体与属于Fournol类群的个体特别接近,这表明该祖先在LGM期间具有局部遗传连续性(补充图)。13).F4-统计数据进一步支持这一观点,揭示了Le Piage II与Fournol星团的关系比vstonice星团更密切(f4(Mbuti, Le Piage II;vstonice, Fournol 85)≫0,Z= 6.58)。我们还比较了它与El Mirón(西班牙北部)的亲缘关系,这是迄今为止测序的最古老的抹大拉人相关个体(19 ka)。F-统计数据表明Le Piage II在基因上介于Fournol 85和El之间Mirón(补充数据2.D).此外,先前的研究表明,El Mirón携带了来自Villabruna聚类的遗传贡献,该聚类在来自意大利的epigravettian相关个体中发现4,15.El Mirón与Villabruna星团的相似性显著高于Fournol 85和Le Piage II,而Le Piage II与Villabruna星团的亲和性并不显著高于Fournol 85(补充数据)2.D).总的来说,与梭鲁特人相关的Le Piage II个体将之前的Fournol祖先与在El Mirón中发现的后续祖先联系起来,为整个西南和西欧LGM的遗传连续性提供了直接证据。因此,这些欧洲地区构成了人类在LGM期间生存下来的气候避难所。
后lgm在意大利半岛
LGM之后,Epigravettian文化在欧洲南部和东南部广泛传播。尽管关于其性质的讨论越来越多27,28在美国,epgravettian文化传统上被认为是从先前的当地Gravettian文化过渡的结果29.然而,与这些文化相关的个体之间的遗传连续性水平以及epigravetan相关个体之间的种群结构尚未得到充分探索。在这里,我们报告了来自4个个体的基因组数据,包括来自意大利东北部(Pradis 1)、意大利西北部(Arene Candide 16)和西西里岛(San Teodoro 2)的3个大约1.3万年前的基因组,以及来自Tagliente 2的全基因组覆盖增加15可追溯至17 ka。
在MDS图中,我们发现所有新报道的和之前报道的epigravettian相关个体都属于Villabruna聚类4(无花果。1 c).一系列的f4-对称性统计证实,所有与epigravettian相关的个体都是支系的,并且与任何本地(Paglicci 12)或非本地祖先(Goyet Q116-1, Kostenki 14, Mal 'ta 1或vstonice)没有过度亲缘关系(补充数据)2.F).此外,没有一个epigravettian相关的个体与南欧的关系比与中东欧gravetettian相关的群体更密切,如所示f4(Mbuti, epigravettian相关个人/团体;vstonice, Paglicci 12),与0一致(补充数据2.G).
接下来,我们研究了意大利半岛上epigravetian相关个体之间的遗传关系,通过基于配对矩阵重建系统发育f2遗传距离(图;3和补充图。9),并用f4-表单中的统计信息f4(Mbuti, Epigravettian A;(补充数据2.E).推断的拓扑结构揭示了与个体年龄无关的系统地理模式。特别是,与所有其他epigravettian相关的个体相比,来自意大利东北部的13 ka Pradis 1个体代表了最基础的谱系,包括来自意大利北部的更古老的Tagliente 2和Villabruna基因组(第1组)。来自意大利西北部(Arene Candide 16),意大利中部(Continenza)和西西里岛的个体属于系统遗传学上更派生的分支(第2组)。在西西里岛,14 ka的Oriente C个体与更年轻但地理上更接近的10 ka Uzzo群体具有更高的亲缘关系30.(n= 2)与几乎同时期来自西西里岛东部的圣特奥多罗2号个体相比。
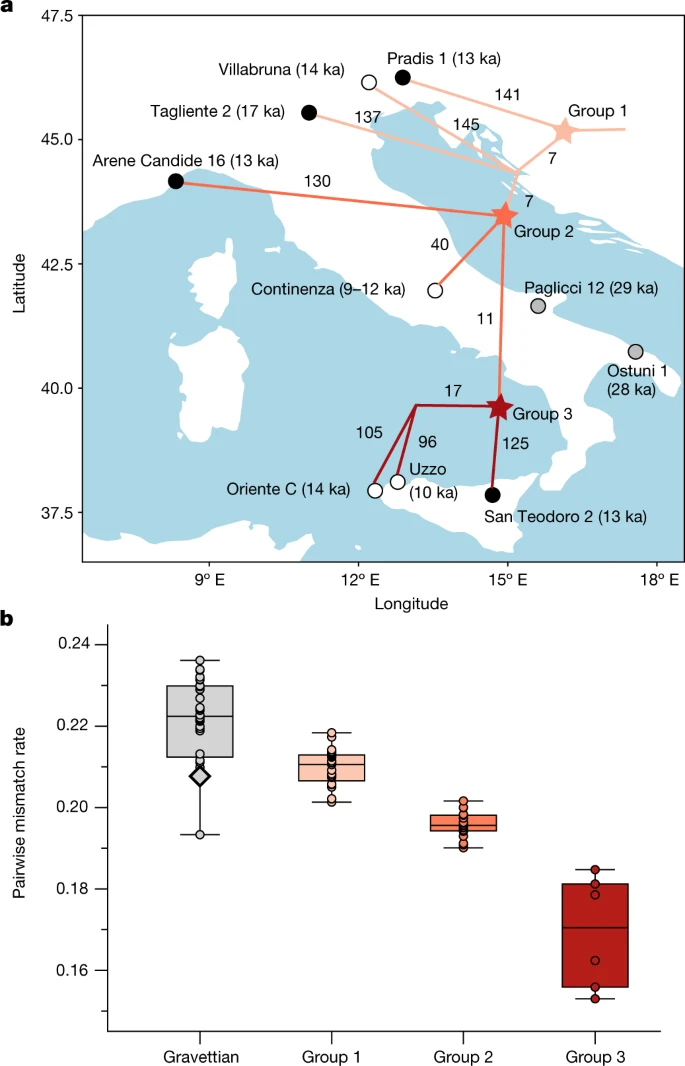
一个,基于两两相邻连接树揭示的epigravettian相关种群的种群结构f2遗传距离。分支标签显示唯一的漂移长度;黑点代表新近获得数据的个体,白点代表先前公布的基因组;每个节点的位置并不意味着分裂发生的位置。b,不同群体个体间PMR表现出的群体多样性。中epigravettian相关种群的分组显示为相同的颜色一个.在Gravettian组中标记出的黑色轮廓钻石显示了来自意大利南部的两个Gravettian相关个体(Paglicci 12和Ostuni 1)之间的PMR。在盒状图中,中线是中位数,盒界描绘了四分位范围,胡须延伸到最大值和最小值,不包括异常值。每组中包含的个体对的样本量在补充数据中报告3.一个.
最后,我们通过计算伪单倍体基因型的配对错配率(PMR)和伪二倍体基因型的个体杂合度水平,估计了数据集中epigravetan相关个体的遗传多样性(补充数据)3.一个).与所有分析的gravettian相关类群中观察到的遗传多样性相比,epigravettian相关个体表现出显著较低的遗传多样性(双尾t以及,P< 0.001)(图3 b).此外,我们发现epigravetan相关群体的遗传多样性从北到南呈下降趋势,意大利北部个体的PMR和杂合度值最高(组1),意大利西部和中部个体的PMR和杂合度值中等(组2),西西里个体的PMR和杂合度值最低(组3)(图3)。3 b).通过分析ROH段,可以观察到类似的模式(扩展数据图。5及补充资料部分9).我们在西西里岛的epigravettian相关个体中检测到最高数量的ROHs,他们携带超过200 cM的短ROHs (4-8 cM)。这表明最近的有效种群规模非常小,估计约为70只(补充表)2),导致西西里Epigravettian狩猎采集者的遗传多样性较低。
总之,我们的研究结果强调了意大利半岛的gravettian- v斯通星系团与epigravettian - Villabruna星系团之间的遗传转换,这可能与考古记录中观察到的不连续有关31.我们表明,所有分析的epigravettian相关个体都具有同质的Villabruna血统,群内遗传结构主要由其地理分布决定,而不是时间分布。epigravettian相关基因组的系统发育重建表明,Pradis 1的分化比其他所有基因组都要深,这表明这种转变发生在17 ka之前——更衍生的Tagliente 2基因组的日期。这与El Mirón 19 ka中Villabruna祖先的证据一起,进一步表明这种遗传不连续可能是与LGM相关的古地理和古生态转变的结果32,而不是b ø ling -艾尔勒ød变暖期4,15(14.7 - -12.9 ka)。此外,我们的系统地理学分析指出,意大利东北部可能是意大利半岛epigravetan相关基因库的入口点。这一发现与Villabruna类群与古代和现代近东祖先的遗传亲和性相结合4,15,16(补充资料,章节8,补充图。18及补充资料2.O),他认为巴尔干半岛是epigravettian相关人口的来源地。因此,LGM可能在阿尔卑斯以南创造了一条从东到西的人类迁徙走廊,从基因上把巴尔干半岛到伊比利亚半岛的狩猎采集者联系起来,也可能是通过沿着现有的低海平面海岸扩散32.
西欧和中欧的后lgm
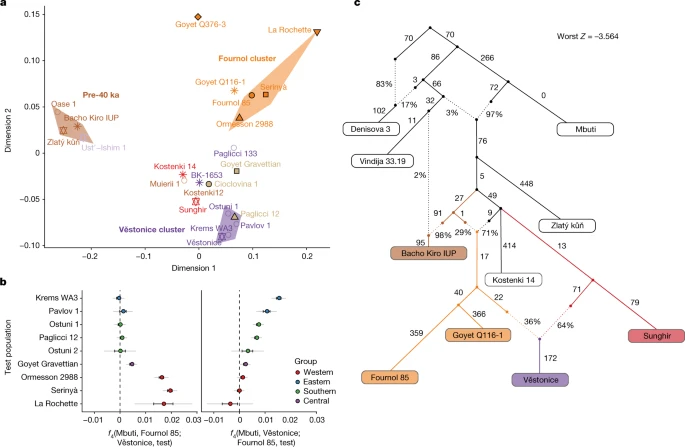
马格达伦文化在LGM之后广泛分布在欧洲西南部、西部和中部33.尽管地理范围很广,但与这种文化有关的不同群体是否起源于一个共同的人群,以及这些群体之间的基因关系如何,目前尚不清楚。先前的研究在与马格达伦人相关的个体中确定了两种不同的遗传组成——包括中欧-西欧基因组的GoyetQ2集群,可追溯到大约15 ka(来自法国、比利时和德国),以及来自西班牙的El Mirón个体的祖先4,14大约从19 ka开始。这两个祖先都携带着与可追溯至35 ka的Goyet Q116-1个体远亲的遗传成分,伊比利亚个体也显示出与Villabruna类群的亲缘关系4,14.通过联合分析之前发表的数据和我们最新报道的与分别来自法国西部和北部La Marche (18 ka)和Pincevent (15 ka)以及波兰南部Maszycka (18 - 16 ka)的马格达伦人相关的基因组相关的基因组,我们确认Goyet Q116-1祖先在所有研究过的马格达伦人相关基因组中都存活了下来,除了在西南和西欧的Gravettian和梭鲁特人相关个体中。1).值得注意的是,与Goyet Q116-1相比,Fournol血统为GoyetQ2集群和El Mirón中发现的遗传成分提供了更好的代理2. h).然而,使用f4-统计数据,我们表明,与Fournol集群相比,所有与magdalenian相关的个体,而不仅仅是El Mirón,都具有villabruna相关的祖先(补充数据2. h).与来自意大利北部的epigravettian相关个体(组1)相比,来自意大利西部和中部以及西西里岛的epigravettian相关个体(组2和组3)的亲和性更强(补充数据)2.F).
因此,我们将属于GoyetQ2簇和El Mirón的个体建模为Fournol 85和Arene Candide 16基因组的混合物,分别代表magdalenan相关群体中的Fournol和Villabruna祖先(图2)。4).除了El Mirón有43%的Villabruna血统外,所有其他与magdalenian相关的个体都有较低的比例(19-29%),因此可以被分配到GoyetQ2聚类(图2)。4及补充资料3.摄氏度).进一步验证了这一点f4-表单统计信息f4(Mbuti, Arene Candide 16;Goyet Q-2, magdalenian相关个体),仅对El Mirón显著阳性,而所有其他测试个体和Goyet Q-2与Arene Candide 16对称相关(补充图。26及补充资料2. h).
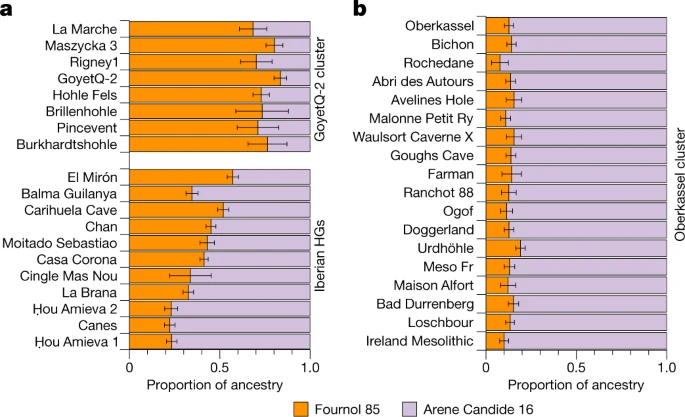
一个,b, GoyetQ-2群和伊比利亚狩猎采集者(HGs)个体的祖先(一个)和Oberkassel集群中的个体(b)使用qpAdm建模,Fournol 85和Arene Candide 16分别代表Fournol和Villabruna祖先。颜色条的长度显示了每个祖先的比例。误差条显示了5厘米分段折刀分析估计的s.e.m.。建模的细节在补充数据中提供3.摄氏度.
我们的分析表明,与Goyet Q116-1相比,Fournol簇是一个更好的magdalenan相关基因组来源。因此,在这些后lgm个体中发现的大多数祖先可能追溯到来自西欧和西南欧洲的gravettian相关群体。与Villabruna祖先的遗传亲缘关系存在于El Mirón和来自西欧和中欧的magdalenian相关个体中。这表明,欧洲南部和西南部狩猎采集者之间的遗传联系在LGM前后延伸到了比利牛斯山脉北部。由此得出的GoyetQ2聚类包括了从法国西部到波兰的18 - 15 ka时期的个体。因此,与之前的建议相反34,这表明后lgm时期的马格达伦扩散确实与西欧向北和东北的人口扩张有关35.
14 ka后至新石器时代
先前的研究表明,在大约14 ka之后,欧洲大部分地区主要有两种主要的狩猎-采集祖先,即西部狩猎-采集(WHG)祖先,与Villabruna类群有关,以及东部狩猎-采集(EHG)祖先,显示出与旧石器时代晚期西伯利亚个体中发现的Villabruna和古代北欧亚(ANE)祖先的亲缘关系4,36.来自北欧和东欧不同地区的携带混合WHG/EHG基因图谱的狩猎采集者已经进行了测序,提出了这两种类型的祖先如何形成并在时间和空间中相互作用的问题37,38,39,40.
在MDS图(图。1 c)和西欧亚主成分分析(PCA)(扩展数据图。6和补充图。14),大多数来自西欧和中欧的14 ka后个体接近WHG类群,来自东欧的个体接近EHG类群,而来自中亚的Tutkaul 1个体接近ane相关类群。这两个14ka的Oberkassel个体标志着阿尔卑斯山以北最早的WHG祖先的存在,因此我们将其重新命名为Oberkassel集群(以下简称Oberkassel集群或祖先),使用迄今为止报道的最古老的个体的名字,携带这种覆盖超过一倍的祖先,以保持一致性4.以…为基础f4-统计数据,我们发现分配到Oberkassel集群的个体比来自意大利的任何其他epigravetian相关群体更接近Arene Candide 16基因组(补充数据)2.F).此外,Oberkassel集群携带Villabruna祖先和GoyetQ2祖先的贡献(补充数据2.J).qpAdm证实了这一点,在qpAdm中,我们可以将所有来自Oberkassel星团的个体建模为一个大致恒定的混合物,大约75%的芳烃Candide 16和25%的Goyet Q-2(或90%的芳烃Candide 16和10%的呋喃85)。4 b及补充资料3.摄氏度).观察到14ka后的人来自西欧和中欧,也来自英国41携带同质的基因组成,而不是显示与GoyetQ2祖先重复的局部混合,这意味着oberkassel祖先的轮廓在其扩散之前已经基本形成。这与伊比利亚狩猎采集者的遗传历史形成了鲜明的对比,在伊比利亚狩猎采集者中,Villabruna/Oberkassel祖先的传播涉及到多次当地与携带高比例GoyetQ2祖先的群体的混合事件14(无花果。4及补充资料3.摄氏度).伊比利亚半岛持久的遗传连续性还体现在y染色体单倍群C的保存直到中石器时代,这在LGM之前的群体中占优势,但在欧洲其他地区的LGM之后很少发现(扩展数据图)。1而且2).
使用f4根据统计数据和qpAdm,我们确认东欧EHG人群是Villabruna/Oberkassel和ANE血统的混合(补充信息,章节11及补充资料2.K).F4-统计数据还显示,来自俄罗斯西部卡累利阿的约8.2 ka Yuzhniy Oleniy Ostrov类群由19个基因组组成,与所有其他EHG类群相比,与Villabruna血统具有相当或更低的亲缘性(补充数据)2.K).与Yuzhniy Oleniy Ostrov族群相比,最古老的个体是来自俄罗斯西部萨马拉的11 ka Sidelkino个体42.为了与前面讨论的命名法保持一致,我们将EHG祖先重命名为Sidelkino群集(以下称为Sidelkino群集或祖先)。Oberkassel和Sidelkino聚类之间的遗传差异在单亲遗传标记的多样性上也很明显,因为Oberkassel聚类以mtDNA单倍群U5和y染色体单倍群I为主导,而来自Sidelkino聚类的个体显示出更高频率的mtDNA单倍群U2, U4和R1b,并且携带唯一的y染色体单倍群Q, R和J(扩展数据图。1而且2).
然后,我们尝试用qpAdm作为Oberkassel、Sidelkino、GoyetQ2祖先的混合,以及安纳托利亚新石器时代农民(ANF)的祖先,对14-5 ka的250个已发表和新报道的狩猎-采集者进行建模,因为相当一部分测序的狩猎-采集者基因组可追溯到大约8 ka之后,当时ANF祖先开始在欧洲传播。我们的结果表明,Oberkassel和Sidelkino祖先之间的接触带和混合模式随着时间的推移而变化(图2)。5).在14到8 ka之间,西欧和中欧的所有狩猎采集者都只有Oberkassel血统,没有发现来自Sidelkino集群的贡献。再往北和往东,来自波罗的海(Baltic HG)、斯堪的纳维亚(SHG)、巴尔干(Iron Gates HG)和乌克兰(Ukraine HG)的人已经拥有了奥伯卡塞尔/Sidelkino混合血统38,408 ka以前。此外,这些群体对ANF也有亲缘关系,这表明在他们的人口统计学历史背后有更复杂的遗传过程16.此外,来自俄罗斯西部的两个最古老的群体属于Sidelkino星系团- peschanitsa (13 ka)43新报告的Minino个体(11 ka)显示出与Oberkassel类群的额外亲缘关系,这可能是由于在sidelkino祖先剖面的初始形成阶段,这种祖先比例的变化。使用DATES软件,我们估计这些老sidelkino集群相关个体的Villabruna/Oberkassel和ANE祖先之间的混合约为15-13 ka(扩展数据图)。7及补充表3.),这与欧伯卡塞尔人祖先在中欧的首次出现大致吻合。这提出了Oberkassel星系团的取代和Sidelkino星系团的形成可能是b ø ling - aller ød间期突然变暖影响人口扩张的结果4,24.
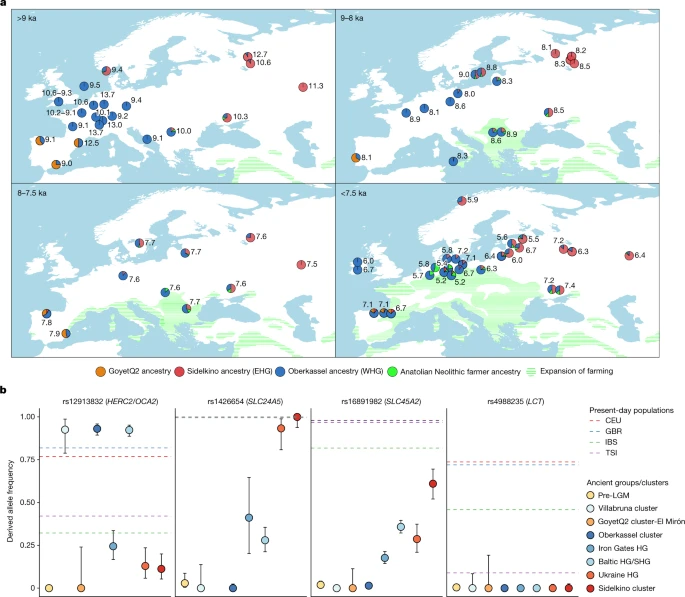
一个使用qpAdm建模了14 ka至5.2 ka之间的狩猎采集者的遗传祖先,其中Oberkassel, Yuzhniy Oleniy Ostrov, Goyet Q-2和来自当今土耳其(barcan, menteye和Boncuklu遗址)的新石器时代农民分别代表Oberkassel (WHG)(蓝色),Sidelkino (EHG)(红色),GoyetQ2(橙色)和安纳托利亚新石器时代农民(绿色)祖先。图中显示了平均校准日期,饼图显示了每个群体或个人的估计祖先比例。建模的细节在补充数据中提供3.E,F.农业扩张9、8、7.5和7 ka用绿色阴影表示。改编自https://doi.org/10.5281/zenodo.5903165(cc乘4.0)。b,不同狩猎采集者群体在与肤色相关的四个snp上的等位基因频率(彩色点)(SLC24A5而且SLC45A2)、眼睛颜色(HERC2/OCA2)和乳糖酶持久性(临床上).点表示最大似然估计,误差条表示衍生等位基因频率的95%置信区间(n,每组的个体数目,载于补充数据3.G).虚线显示了目前1000个基因组计划人群的估计频率(CEU,北部和西欧血统的犹他州居民;GBR,英国;肠易激综合症、西班牙语;托斯卡纳TSI)37.有关等位基因频率估计的详细信息见补充信息,部分12,补充图。27而且28及补充资料3.G.
从大约8 ka开始,我们开始观察中欧与Sidelkino血统的混合事件。这首先在德国东北部的Gross Fredenwalde的个体中被检测到,此后在大多数欧洲狩猎采集个体中达到约10%(扩展数据图)。8).8 ka后不久,Sidelkino祖先在西班牙东部消失了,但随着Oberkassel祖先的增加,它已经到达了伊比利亚北部。5).相反,在东欧,来自上伏尔加河地区的米尼诺I和雅兹科沃人新生成的基因组中,至少有7.5 ka的奥伯卡塞尔血统被鉴定出来,而来自米尼诺I的1000岁以上的个体则没有这种遗传成分。考虑到伏尔加河上游地区的淡水水库信号,人类遗骸的放射性碳年代看起来比他们的真实年龄要老500年左右44在美国,西德基诺血统的中欧狩猎采集者与奥伯卡塞尔血统的东欧狩猎采集者之间可能存在1000多年的时间间隔。然而,需要在时间和空间上进行额外的基因组中间,以评估这两种混合事件是独立的还是共同人口统计过程的一部分。
7.5 ka后,随着ANF祖先到达阿尔卑斯山以北地区,携带狩猎-采集遗传特征的个体主要局限于欧洲北部边缘地区(图2)。5).在这一时期,奥伯卡塞尔血统的混合进一步向东扩散,大约6.5 ka到达萨马拉,并且在波罗的海地区的狩猎采集者中发现了Sidelkino血统的增加,这之前与纳尔瓦文化向梳状陶瓷文化的过渡有关38,39(扩展数据图。8).在中欧,与ANF血统的混合变得非常普遍,但并不是无处不在,这表明狩猎采集社会和农民社会共存了几百年,没有混合。在分析的数据集中,最年轻的携带大量狩猎-采集祖先的个体来自德国北部的Ostorf,可追溯到大约5.2 ka (>90% Oberkassel聚类加上sidelkino聚类成分)(补充数据3.F)。在这个遗址上发现的个体可能标志着如此高水平的与狩猎采集相关的祖先的最后一次出现,就在欧洲青铜时代出现的几个世纪之前。
基于主成分分析和外群分析f3.根据统计数据,来自塔吉克斯坦的新石器时代的图特考尔1号人与来自西伯利亚中南部的旧石器时代晚期的人(Afontova Gora 3号(AG3)和Mal 'ta 1号),以及大约同时期的西西伯利亚狩猎采集者(秋明和Sosnoviy)密切相关,两者都有很高比例的ANE血统45(无花果。1 c和扩展数据图。6).我们测试了Tutkaul 1相对于AG3与世界古代和现代种群的亲缘关系。与西西伯利亚的狩猎采集者相反,图考尔1号没有额外的欧亚大陆东部血统,但与伊朗新石器时代的农民和伊朗和图兰地区的一些年轻人口有亲缘关系(补充数据)2.l).相反,Sidelkino集群中的个体在基因上更接近AG3而不是Tutkaul 1。这表明,新报道的来自中亚的新石器时代个体的祖先可能是伊朗和图兰地区与ane相关贡献的一个很好的代表45大约5.5 ka,但不是来自东欧的狩猎采集者。
总之,我们描述了14 ka以来欧洲两个主要的狩猎采集祖先Oberkassel和Sidelkino集群之间的形成和相互作用。Oberkassel星团与意大利西北部的Arene Candide 16的基因组相似性可能意味着epigravetan相关的祖先从欧洲南部通过阿尔卑斯山地区的西侧传播到中欧。Sidelkino血统也在14ka左右出现,在东欧有第一个直接证据43追溯到13 ka。从大约8 ka开始,不同的狩猎-采集群体之间的混合程度不断增加,表明这些觅食群体的流动性增强了。这在一定程度上可能是由新石器时代农民在欧洲的扩张和/或环境因素引发的,例如8.2 ka左右的气候事件,这是全新世时期北半球最大的突然冷却46,47.
表型相关变异
利用大量增加的样本量,我们调查了遗传上不同的狩猎-采集群体,在已知与现代欧洲人特定表型性状相关的特定位点上的等位基因频率。5 b和补充图。27而且28).与之前的发现一致,所有被分析的组都没有显示出在SNP rs4988235上的衍生等位基因临床上基因,负责乳糖酶的持久性。如前所述37,我们发现在lgm后的狩猎-采集群体中,与皮肤和眼睛色素沉着相关的等位基因频率变化很大。对于与浅眼睛颜色相关的SNP (HERC2 / OCA2(rs12913832)),来自Villabruna聚类、Oberkassel聚类、Baltic HG和SHG组的个体表现出高频率的衍生等位基因(>90%),这是导致绿色或蓝色眼睛表型的原因,而Sidelkino聚类、乌克兰HG和Iron Gates HG组的该等位基因发生率较低(10-25%)。相反,对于与肤色相关的两个snp (SLC24A5(rs1426654)和SLC45A2(rs16891982))、Sidelkino聚类和Ukraine HG群的频率较高(>90%)SLC24A529-61%的人SLC45A2与Oberkassel和Villabruna聚类相比,这些等位基因几乎完全不存在(<1%)。根据现代欧洲人的遗传变异,这可能意味着14 ka后欧洲各地狩猎-采集人群之间的表型差异,Oberkassel集群中的个体可能表现出较深的皮肤和较浅的眼睛,Sidelkino集群中的个体可能表现出较浅的皮肤和较深的眼睛颜色。
讨论与结论
本研究中产生的数据使我们能够在高分辨率下研究欧亚狩猎-采集者之间的基因组转换和相互作用(扩展数据图)。9).我们对从旧石器时代晚期到新石器时代的30000年时间跨度内狩猎-采集人群的基因组历史提供了五个新的见解。
首先,我们表明,在整个欧洲,与Gravettian文化相关的个体在生物学上并不是一个同质的种群。然而,在文化上,我们看到了广泛的总体趋势,如武器和一些可携带的艺术48,以及其他更具区域特色的方面,例如殓房惯例(补充资料,第13),各种新颖的岩石和硬有机材料工具包和装饰品20.,21.在中欧的Aurignacian文化相关的个体中发现的祖先(GoyetQ116-1祖先)产生了来自西欧和西南欧洲的gravetian相关的个体。这个衍生的祖先- Fournol集群-在LGM期间在梭鲁特人相关的个体中幸存下来,可能在法国-坎塔布安气候避难区25,导致后来与Magdalenian文化相关的人口(GoyetQ2集群和El Mirón)。相反,在30 ka前东欧个体(Kostenki类群和Sunghir类群)中发现的祖先有助于中欧和南欧的gravettian相关个体(vstonice类群),后者没有从这些地区的lgm后种群中提取的后代。
第二,与Epigravettian文化(Villabruna集群)相关的个体的祖先,被发现在基因上连接了欧洲和近东的狩猎-采集者,在早期和晚期Epigravettian之间的过渡之前很久就到达了南欧4,15而且可能早在格拉维-埃格拉维过渡时期。对携带这一祖先的不同血统的系统地理学重建进一步表明,它的入口点从巴尔干半岛进入意大利东北部,然后从北到南扩展到意大利半岛,同时由于连续的瓶颈,人口下降。
第三,与magdalenian相关的个体不仅来自伊比利亚,而且来自欧洲其他地区,都带有epigravetian相关的祖先(Villabruna集群)。对与先前巴德古连文化有关的西欧个体的遗传分析可能为导致GoyetQ2类群形成的过程提供线索。根据考古记录推断35在美国,马格达伦人在欧洲的传播与lgm后西南到北部和东北部的人口扩张有关,而不是来自东南部难民的迁移34.
第四,我们扩展了早在14 ka中欧和西欧狩猎采集者中发现的大规模基因更替,这些人与多个技术综合体有关——federmesser, Azilian和其他最终旧石器时代群体4——尽管在技术上与之前的马格达伦晚期有相当大的连续性。这个分布广泛的祖先(Oberkassel群集(也称为WHG))与来自意大利西北部的epigravetan相关个体关系最密切,这表明它向欧洲大陆的扩张可能是从阿尔卑斯山的西边开始的,而不是东边。此外,与magdalenian相关的基因库几乎完全的基因替换提出了这样一个假设,即在大约14.7 ka开始的b ø ling - allerd变暖期的突然气候变化期间,欧洲部分地区的人口分布有所不同,从而创造了南欧人口可以扩张的地区。这可能也解释了横跨欧亚大陆西部大部分地区的奥伯卡塞尔星群的遗传一致性,但需要15至14 ka的基因组数据才能了解这种更替的确切动态。
第五,西欧和中欧的Oberkassel祖先和东欧的Sidelkino祖先在近6000年的时间里基本上是孤立的,直到基因相互作用首次被观察到——在德国东北部大约8 ka,可能与波罗的海沿岸的文化交流有关49在伏尔加河上游地区约7.5 ka,这可能与该地区陶器的传播有关50.
总之,我们的研究表明,在最后一个冰河时代最冷的阶段,西欧和西南欧洲是人类群体持续生存的气候避难所,而意大利半岛和东欧平原的人口在基因上发生了颠覆,挑战了这些地区作为人类冰川避难所的角色。新来的维拉布卢纳祖先后来成为欧洲最广泛的狩猎采集祖先。对于来自巴尔干半岛的旧石器时代晚期个体的进一步古基因组研究,对于理解东南欧是否代表Villabruna祖先的来源和LGM期间人类人口的气候避难所至关重要。
证明中增加的说明:配套论文51描述了一个来自南伊比利亚的2.3万年前梭鲁特人相关个体的全基因组数据,这些数据扩展了欧洲西南部LGM遗传连续性的证据。
方法
考古抽样
这项研究中分析的古人类标本来自多项科学合作。在负责考古材料分析的机构的批准下,对所有遗骸进行了取样。这是通过与来自骨骼材料保存国家的当地策展人和科学家合作实现的,他们被列入本研究的作者之列。来自每个考古遗址的材料的负责共同作者在补充信息部分列出1.
所分析的个体跨度从旧石器时代晚期到新石器时代。虽然诸如石器工业、技术综合体、史前传统等术语可能更适合指各种相关的时间文化细分,但它们涉及不同的讨论层次,并不适用于本文调查的所有上下文。因此,这里使用的更广泛的术语“考古文化”或简单的“文化”是指考古学定义的物质文化,而不暗示与现代人类学和/或民族志文化概念的联系。
放射性碳年代测定法
我们报告了在曼海姆的Curt-Engelhorn-Zentrum Archaeometrie对40个人的骨骼元素进行的47个新的放射性碳测年(MAMS,n= 29),格罗宁根大学同位素研究中心(GrA和GrM,n= 5),奥胡斯大学(AAR,n= 3), Beta分析(Beta,n= 2), Zürich (ETH,n= 3),国际化学分析(ICA,n= 2),巴黎自然历史博物馆(Echo Lab,n= 1)和维尔纽斯(FTMC,n= 2)(补充数据1.一个).使用OxCal 4.4校准日期52校正曲线IntCal20,概率为95.4%53当同一个人有多个日期时,我们使用函数R_Combine来组合它们52.我们没有修正海洋或淡水储层效应的校准日期,但在可用时,我们报告了单个稳定同位素值(δ15N /δ13C和C:N比值)1.一个评估这种储层效应的潜在影响。
古代DNA处理
人类遗骸在耶拿(德国)的马克斯·普朗克人类历史科学研究所、Tübingen大学(德国)、佛罗伦萨大学(意大利)、莱顿大学医学中心(荷兰)和塔尔图大学(爱沙尼亚)的专门实验室进行处理。在洁净室设施中对人类骨骼和牙齿进行采样,以尽量减少在此过程中现代人的DNA污染。按照既定程序从生成的骨头或牙粉中提取DNA。对样本的子集(GER002和GER003)进行预处理,以减少表面污染54.阴性对照和阳性对照也包括在内。DNA裂解用EDTA 900 μl、H 75 μl溶液2添加O和25 μl蛋白酶K。在旋转器中,样品在37°C下消化至少16小时,然后对预处理样品进行消化55在56°C下再延长一小时。然后将悬浮液离心并转移到前面所述的结合缓冲液中56.为了结合DNA,使用高容量硅胶柱(高纯病毒核酸大容量试剂盒(Roche))。使用制造商的洗涤缓冲液洗涤2步后,DNA在TET (10 mM Tris, 1 mM EDTA和0.05% Tween)中分两步洗脱,最终体积为100 μl。DNA裂解后,在自动液体处理系统(Agilent Technologies Bravo NGS工作站)上使用硅涂层磁颗粒提取样本子集。57.用25 μl DNA提取物构建双链DNA库,不含尿嘧啶DNA糖基酶(ds_nonUDG)或存在尿嘧啶DNA糖基酶(ds_halfUDG),然后用双链“UDG-half”库制剂来减少但不消除古DNA (aDNA)片段末端脱氨诱导的损伤量58.每次实验同时进行阴性和阳性对照。使用IS7和IS8引物对文库进行定量59使用DyNAmo SYBR Green qPCR试剂盒(赛默飞世尔科学公司)对LightCycler 480(罗氏公司)进行定量分析。每个aDNA库都被双索引60使用PfuTurbo DNA聚合酶(安捷伦)进行1-4平行100 μl反应。每个文库的索引产物经MinElute色谱柱(Qiagen)聚合,用50 μl TET洗脱,再用IS5和IS6引物定量59使用上述定量方法。使用Herculase II Fusion DNA聚合酶(Agilent),按照制造商的规格,使用0.3 μM的IS5/IS6引物,在多个100 μl反应中扩增纯化的产物。再次MinElute纯化后,使用安捷伦2100 Bioanalyzer DNA 1000芯片对产品进行量化。然后制备所有文库的等量池,在Illumina Hiseq4000平台上使用75bp单端reads进行测序筛选。在没有尿嘧啶DNA糖基酶(ss_nonUDG)的情况下,用30 μl DNA提取物建立单链DNA文库,然后进行双索引,使用中描述的协议的自动化版本61关于前面提到的液体处理系统。Cuiry Les Chaudardes 1的单链库通过部分UDG处理产生(ss_halfUDG)62(补充数据1.B).
DNA富集和测序
用IS5/IS6引物进一步扩增双链和单链文库,使其在NanoDrop分光光度计(赛默飞世尔科学公司)测得的浓度达到200-400 ng/μl。这些文库在Illumina HiSeq 4000仪器上进行浅散弹枪测序,使用制造商的方案进行75个单端运行周期,以评估人类内源性DNA含量和质量。在鸟枪数据中,人类DNA百分比约为0.1%或更高的样本富集了一组1,237,207个靶向snp(捕获1240k),分布在人类基因组中6.mtDNA捕获63也对那些mtDNA覆盖率不够高的文库进行了检测,以评估mtDNA单倍群和污染。Illumina测序平台也用于测序1240k和mtDNA捕获文库(补充数据1.B).
使用EAGER管道1.92.55对分离多路捕获测序reads进行清理,并将其映射到人类参考基因组hs37d564.在管道中,适配器被AdapterRemoval 2.2.0删除65, reads采用BWA 0.7.12 aln/samse算法进行映射66,使用DeDup 0.12.1 (https://github.com/apeltzer/DeDup),并使用mapDamage 2.0.6和2.0.9检查每个库的损坏模式67.使用PMDtools 0.60过滤重复数据删除的bam文件68阈值为3,以减少基于死后DNA脱氨的潜在现代DNA污染。对于ds_halfUDG库,我们在bamUtil 1.0.13中使用trimBam从读取的两端屏蔽了2 bp (https://github.com/statgen/bamUtil),移走受损地点。
使用AdapterRemoval 2.2.0清洗线粒体捕获测序reads,去除长度低于30 bp的适配器和reads。然后将清洗后的reads与1240k捕获测序的reads一起映射到人类参考线粒体序列NC_012920.1使用BWA 0.7.12 aln/samse算法(参数-n 0.01, -l 16500),并使用CircularMapper重新对齐64.来自同一个人和库设置的映射读取被合并,重复被DeDup删除。然后使用samtools对映射质量低于30的Reads进行过滤,并使用Schmutzi生成共识序列69.
古代DNA鉴定和基因分型
每个个体的性别由性染色体和常染色体的测序覆盖率的比例决定(补充数据1.摄氏度).使用pmd过滤的bam文件对库显示受污染迹象的个体进行进一步测试。至少有一个库显示Y/Auto比值> 0.2的个体被判定为男性个体,Y/Auto < 0.2的个体被判定为女性个体4(补充图。12).
用几种方法估计了核DNA污染。我们应用ANGSD 0.93470和hapCon71男性个体的文库和应用污染72以及一种新开发的分析女性和男性图书馆ROH污染的方法(见补充信息,第2以获取详细描述)。mtDNA污染用Schmutzi(——notusepredC——uselth)估计。69所有的库。显示线粒体或核污染率超过10%的图书馆被认为是严重污染,而介于5%至10%之间的图书馆被认为是轻微污染,并被区别对待(详细信息见补充信息部分)3.).
在SAMtools 1.3中使用mpeup对基本质量和映射质量大于30的清理后的读进行堆积73在1240k个目标站点上。对于污染的文库,我们使用pmd过滤的bam文件作为基因分型的输入。伪单倍体基因型采用pileupCaller 1.4.0.2 (https://github.com/stschiff/sequenceTools)在随机单倍体调用模式下。对于ds_halfUDG库,我们将2bp-mask bam文件中所有目标位点的基因型称为基因型;对于ds_nonUDG文库,我们只在转化位点上称其为基因型;对于ss_nonUDG文库,我们称之为单链模式的基因型,这种模式忽略了C/T多态性的正向读和G/A多态性的反向读。
然后,我们使用自定义脚本,从可用的基因型调用中随机选择等位基因,合并同一个体的不同基因型库。合并后,由于低覆盖率,在1240k个位点上snp小于6000个的个体被排除在进一步分析之外。我们也用同样的方法选择了先前发表的个体进行基因分型(补充数据1.G)2,4,30.,74,75,76,77,78.然后,我们将我们新生成的基因型与AADR v42.4中发表的古代和现代个体的基因型相结合。https://reich.hms.harvard.edu/allen-ancient-dna-resource-aadr-downloadable-genotypes-present-day-and-ancient-dna-data)版本42.4)进行下游分析1,4,7,14,16,23,36,37,38,39,40,42,43,45,79,80,81,82,83,84,85,86,87,88,89,90,91,92,93,94.
在计算个体杂合度时,我们也从每个文库调用伪二倍体基因型,使用pileupCaller 1.4.0.3在随机二倍体调用模式下,对不同类型文库采用与伪单倍体基因型调用相同的策略。
单性生殖的标记
线粒体单倍群用HaploGrep 2进行测定95,基于Schmutzi生成的一致性序列,在增加质量过滤器(从q0到q20)下对每个样本进行检查。不一致的单倍群分配被手动验证24(补充数据1.l).系统发育重建(扩展数据图。1我们使用了MUSCLE (- maxisters 2)96根据定义的阈值(最小平均覆盖率>5倍,污染估计<20%,HaploGrep 2单倍群分配与手动分配一致),创建先前发表的序列和新报道的mtDNA一致序列的多基因组比对。我们从相同位置移除小于6.5 ka的个体和mtDNA后,构建了具有103个mtDNA序列的Maximum Parsimony树,加上一个非洲序列作为外群(未显示)。该树在16,528个位置(部分删除95%)上计算,并使用MEGA10进行500次自举迭代97.
为了确定男性个体的y染色体单倍群,我们使用来自国际遗传谱系学会(ISOGG)数据集的Y-SNP列表(v.15.73)对y染色体读数进行基因分型,分别忽略正向读数和反向读数上的C-to-T和G-to-A跃迁。这个过程允许我们手动遍历ISOGG y单倍群树,以半自动的方式检查哪些位置被覆盖。这一过程允许我们为覆盖的分支分配一个祖先或派生的单倍群,并在某些情况下对调用进行修正,例如,由于诊断snp末端读位置的残余古代损伤(C-to-T或G-to-A错配)而调用一个更派生的单倍群98(补充数据1.一个).对于个体在y染色体系统发育树上的位置(扩展数据图。1),我们使用pathPhynder99基于卡尔明等人的树。One hundred..我们使用默认的后验阈值0.01,映射和测序质量截断值为30。然后,我们删除了小于0.04X覆盖率的样本(在Y染色体的可映射、非重组区域上计算)98),以避免在主要单倍群的根位置任意放置低覆盖率的样本。这导致了一个有57个新报道和先前发表的古代个体的树,而现代序列在主要的y染色体单倍群中被折叠(最基础的谱系没有显示出来)。基于单倍群分配的低覆盖率古代个体的暂定位置(补充数据)1.一个)在各自的分支上用箭头表示。
生物亲缘性和种群多样性
通过计算亲缘系数(r)基于常染色体SNPs的PMR(补充数据1.F及补充资料部分4).利用伪二倍体基因型估计的个体平均杂合率确定每个群体的基线(补充数据)1.E).
利用hapROH鉴定了狩猎-采集基因组中的ROH片段101.根据建议,我们分析了1240k面板上超过400,000个SNPs的个体101我们称ROH长于4cm(补充数据3.B).有效人口规模(Ne),然后使用最大似然方法,在过滤具有近亲近亲繁殖信号的个体(其基因组在ROH中至少有50厘米的个体,范围为> - 20厘米)(补充信息,节)9).
群体遗传分析
采用EIGENSOFT 6.0.1中的smartpca进行主成分分析102,用现代个体进行计算,所有古代个体都投影在计算后的电脑上。使用“lsqproject: YES”参数来最小化远古个体数据缺失的影响。在1240k_HO数据集上计算了来自87个欧亚西部现代人群的1379个个体,该数据集与1240k数据集和人类起源数据集相交(补充数据)1.K).
MDS分析显示了欧洲狩猎采集者之间的遗传亲和性,该分析是基于来自外群体的距离矩阵f3.-statistics,格式为1−f3.(Mbuti.DG;pop1, pop2),并使用r3.5.1中实现的经典MDS算法(cmdscale)执行。狩猎-采集者根据他们的地理起源和日期分组(补充信息,章节5).的f3.-统计数据是用ADMIXTOOLS 5.1包中的qp3Pop 435计算的103.
成对的f2使用ADMIXTOOLS 7.0.2包中的qpfstats 200生成epigravetan相关基团的距离矩阵,参数为“allsnps: YES, scale: NO”,Mbuti。DG设为外组。然后利用Ape 5.3包中实现的邻域连接方法重构邻域连接树104R 3.5.1。
的f4-统计数据由qpDstat 755计算,参数为“f4 mode: YES”,使用Mbuti。来自非洲的DG人口被全部用作外群体f统计分析。应用ADMIXTOOLS 5.1中的qpAdm 810工具对混合种群的祖先进行建模,“allsnps”模式和补充信息部分中描述的外群集选择11.使用qpGraph 6450重建掺合图,采用allsnps模式对低覆盖率样本和Mbuti进行校正。DG设为外组。使用基于祖先协方差模式的753年日期程序测定外加剂事件的日期105,以0.1 cM为仓大小进行协方差计算,指数拟合开始于d≥0.5厘米。
表型SNP分析
由于大多数古代样本的覆盖率不足以进行二倍体基因型调用,我们计算了涵盖参考或替代等位基因上所选表型snp的reads,并按照Mathieson等人描述的最大似能方法计算了基于组的等位基因频率。37.关于参与分析、读计数处理和等位基因频率计算的个体的详细信息在补充信息部分提供12及补充资料1.J而且3.G.
报告总结
有关研究设计的进一步资料,请参阅自然组合报告摘要链接到这篇文章。
数据可用性
本研究中报告的所有具有新基因组数据的个体的对齐序列可在欧洲核苷酸档案(ENA)上获得PRJEB51862.已编译的用于分析的基因型文件,包括重新基因分型的已发表基因组,已上传至马克斯普朗克学会埃德蒙数据存储库(https://edmond.mpdl.mpg.de/dataset.xhtml?persistentId=doi:10.17617/3.Y1KJMF).
代码的可用性
新开发的基于ROH的污染估计方法的代码可在https://github.com/hyl317/hapROH.用户手册包括安装指南可在https://haproh.readthedocs.io/en/latest/hapROH_with_contamination.html.用于此工作的版本存档在https://edmond.mpdl.mpg.de/dataset.xhtml?persistentId=doi:10.17617/3.Y1KJMF.
参考文献
Prüfer, K.等。来自捷克Zlatý kzheven的一个距今超过45000年的现代人类头骨的基因组序列。Nat,生态。另一个星球。5, 820-825(2021)。
Hajdinjak, M.等人。最初的旧石器时代晚期的欧洲人有尼安德特人的祖先。自然592, 253-257(2021)。
奥拉尔德,I. & Posth, C.西欧亚人考古研究的最新趋势。咕咕叫。当今。麝猫。Dev。62, 36-43(2020)。
傅,Q.等。冰河时代欧洲的遗传史。自然534, 200-205(2016)。
格林,R. E.等。尼安德特人基因组的草图序列。科学328, 710-722(2010)。
傅,Q.等。尼安德特人来自罗马尼亚的早期现代人,其祖先是尼安德特人。自然524, 216-219(2015)。
傅,Q.等。来自西伯利亚西部的4.5万年前现代人的基因组序列。自然514, 445-449(2014)。
Seguin-Orlando, A.等。欧洲人的基因组结构可以追溯到至少36200年前。科学346, 1113-1118(2014)。
Vallini, L.等。遗传和物质文化支持了从非洲人口中心向旧石器时代欧亚大陆的反复扩张。基因组医学杂志。另一个星球。14,疏散045(2022)。
Maier, A. & Zimmermann, A.人口南下?从古人口学的角度来看格拉维特人。古代91, 573-588(2017)。
杜鲁哈诺夫,P. in东欧的文化转型与互动(eds Chapman, J. & Dolukhanov, P.) 122-145 (Avebury, 1993)。
甘伯,C.,戴维斯,W.,佩蒂特,P.,黑兹尔伍德,L.和理查兹,M.晚期冰期欧洲人口的考古和遗传基础:对“农业思维”的影响。剑桥Archaeol。J。15, 193-223(2005)。
Wren, C. D. & Burke, A.末次冰期(LGM)西欧人类种群的生境适宜性和遗传结构。《公共科学图书馆•综合》14, e0217996(2019)。
Villalba-Mouco, V.等人。伊比利亚半岛晚更新世狩猎采集祖先的生存。咕咕叫。医学杂志。29, 1169 - 1177。e7(2019)。
Bortolini, E.等。早期的阿尔卑斯占领可以追溯到晚期欧洲的人类向西迁徙。咕咕叫。医学杂志。31, 2484 - 2493。e7(2021)。
费尔德曼等人。更新世晚期的人类基因组表明,安纳托利亚中部的第一批农民起源于当地。Commun Nat。10, 1218(2019)。
Petr, M., Pääbo, S., Kelso, J. & Vernot, B.长期选择对尼安德特人遗传的限制。国家科学院学报美国116, 1639-1644(2019)。
哈里斯,K. &尼尔森,R.尼安德特人渗入的遗传成本。遗传学203, 881-891(2016)。
《格拉韦特人的起源》。皮疹。Int。359, 3-18(2015)。
古塔斯,nLes Gravettiens(编者Otte, M.) 105-160 (Errance, 2013)。
Klaric, L., Goutas, N., Laccarière, J. & Banks, W. E. in .Les Sociétés Gravettiennes du Nord-Ouest Européen: Nouveaux Sites, Nouvelles Données, Nouvelles Lectures(eds Touzé, O., Goutas, N., Salomon, H. & Noiret, P.) 323-266 (press Univ. de Liège, 2021)。
Mounier, A.等人。欧洲旧石器时代晚期的Gravettian颅骨形态与人类群体亲和性。科学。代表。10, 21931(2020)。
西科拉等人。古代基因组显示了旧石器时代早期觅食者的社会和生殖行为。科学358, 659-662(2017)。
波斯特,C.等。更新世线粒体基因组表明,非非洲人的一次主要分散和欧洲晚冰期的人口周转。咕咕叫。医学杂志。26, 827-833(2016)。
最后一次极寒期人类对欧洲西南部的占领:法国和伊比利亚的梭鲁特文化适应。j . Anthropol。Res。71, 465-492(2015)。
Lécuyer, C., hillire - marcel, C., Burke, A., Julien, M. A. & Hélie, J. F.西南欧洲LGM人类避难所的温度和降水体制从δ推断13C和δ18大型哺乳动物遗骸的O。皮疹。科学。牧师。255, 106796(2021)。
jindjian, F.欧洲末次冰期末期狩猎-采集群体的领土和经济。皮疹。Int。412, 37-43(2016)。
Ruiz-Redondo等人。亚得里亚海盆地旧石器时代中期和晚期:年代学,过渡和人类对不断变化的景观的适应。皮疹。科学。牧师。276, 107319(2022)。
拉普拉斯,G。埃塞德类型学Systématique(Annali dell 'Università di Ferrara, 1964)。
Yu, H.等。西西里岛中石器时代和新石器时代的基因组和饮食不连续性。iScience25, 104244(2022)。
帕尔马迪切斯诺拉,A。Le paléolithique supérieur en Italie。联赛”Prehistoire d 'Europe”9(版本,2001)。
Peresani, M.等。末次盛冰期亚得里亚海波河地区的狩猎采集者:环境和文化动态。皮疹。Int。581 - 582, 128-163(2021)。
马格达伦文明的出现、扩张和稀释。皮疹。Int。272 - 273, 354-361(2012)。
迈尔,A.在中欧的马格达伦人81 - 180https://doi.org/10.1007/978-94-017-7206-8_6(施普林格,2015)。
kozowski, s.k, pouttowicz -Bobak, M., Bobak, D. & Terberger, T.来自Maszycka洞穴和中欧晚冰期再殖民的新信息。皮疹。Int。272, 288-296(2012)。
Raghavan等人。旧石器时代晚期西伯利亚基因组揭示了美洲原住民的双重祖先。自然505, 87-91(2014)。
马西森等人。230个古代欧亚人的全基因组选择模式。自然528, 499-503(2015)。
马西森等人。东南欧的基因组史。自然555, 197-203(2018)。
Mittnik, A.等人。波罗的海地区的遗传史前。Commun Nat。9, 442(2018)。
Günther, T.等。中石器时代斯堪的纳维亚的人口基因组学:研究早期冰川后迁移路线和高纬度适应。公共科学图书馆杂志。16, e2003703(2018)。
查尔顿,S.等。英国晚冰期旧石器时代的双重祖先与生态。Nat,生态。另一个星球。6, 1658-1668(2022)。
Damgaard, P.等人。第一批马牧民和青铜器时代早期草原向亚洲扩张的影响。科学360, eaar7711(2018)。
Saag, L.等人。东欧平原石器时代向青铜器时代过渡过程中遗传祖先的变化。科学。睡觉。7, eabd6535(2021)。
伍德,R. E.等。俄罗斯西北部Minino墓地的淡水放射性碳储层效应。放射性碳测定年代55, 163-177(2013)。
Narasimhan, v.m.等人。南亚和中亚人类种群的形成。科学365, eaat7487(2019)。
格罗宁伯恩,D. in新石器时代向中欧的传播(RGZM, 2010)。
欧洲分子生物地理学:更新世旋回和冰后趋势。前面。黑旋风。4, 11(2007)。
Roebroeks, W., Mussi, M., Svoboda, J. & Fennema, K.。黄金时代的猎人:欧亚大陆旧石器时代中晚期,3万-2万bp(莱顿大学,2000)。
Kotula, A, Piezonka, H. & Tergerger, T. Groß Fredenwalde(德国东北部)的中石器时代墓地及其文化从属关系。Liet。Archeol。46, 65-84(2020)。
Piezonka, H.等人。乌拉尔和西西伯利亚狩猎采集陶器的出现:新的年代测定和稳定同位素证据。j . Archaeol。科学。116, 105100(2020)。
Villalba-Mouco, V.等人。一名2.3万年前的南伊比利亚人将末次冰期前后生活在西欧的人类群体联系在一起。Nat,生态。另一个星球。https://doi.org/10.1038/s41559-023-01987-0(2023)
Bronk Ramsey, C.放射性碳年代的贝叶斯分析。放射性碳测定年代51, 337-360(2009)。
雷默,P. J.等人。IntCal20北半球放射性碳年龄校准曲线(0-55 cal k英国石油公司).放射性碳测定年代62, 725-757(2020)。
科列维奇,P.等人。减少古代骨骼和牙齿DNA提取中的微生物和人类污染。生物学技术59, 87-93(2015)。
Rohland, N. & Hofreiter, M.从骨头和牙齿中提取古代DNA。Protoc Nat。2, 1756-1762(2007)。
Dabney, J.等人。由超短DNA片段重建的中更新世洞穴熊的完整线粒体基因组序列。国家科学院学报美国110, 15758-63(2013)。
Rohland, N, gllocke, I, Aximu-Petri, A. & Meyer, M.从古代骨骼、牙齿和沉积物中提取高度降解的DNA用于高通量测序。Protoc Nat。13, 2447-2461(2018)。
Rohland, N., Harney, E., Mallick, S., Nordenfelt, S. & Reich, D.部分尿嘧啶- DNA -糖基化酶筛选古代DNA的处理。费罗斯。反式。r . Soc。B370, 20130624(2015)。
Meyer, M. & Kircher, M. Illumina测序库制备高复用目标捕获和测序。冷泉港。Protoc。2010, pdb。prot5448(2010)。
Kircher, M., Sawyer, S. & Meyer, M.双索引克服了Illumina平台上多路测序的不准确性。核酸测定。40, e3(2012)。
Gansauge, M., Aximu-Petri, A., Nagel, S. & Meyer, M.单链DNA文库的手工和自动制备,用于古代生物遗骸和其他高度降解DNA来源的DNA测序。Protoc Nat。15, 2279-2300(2020)。
梅耶等人。来自古代丹尼索瓦人的高覆盖率基因组序列。科学338, 222-226(2012)。
傅,Q.等。中国天元洞早期现代人的DNA分析。国家科学院学报美国110, 2223-2227(2013)。
Peltzer, A.等。EAGER:高效的古代基因组重建。基因组医学杂志。17, 60(2016)。
Schubert, M., Lindgreen, S. & Orlando, L. AdapterRemoval v2:快速适配器修剪,识别和读取合并。BMC Res. Notes9, 88(2016)。
李,H. & Durbin R.快速和准确的短读对齐与Burrows-Wheeler变换。生物信息学25, 1754-1760(2009)。
Jónsson, H., Ginolhac, A., Schubert, M. Johnson, P. L. F. & Orlando, L. MapDamage2.0:古代DNA损伤参数的快速近似贝叶斯估计。生物信息学29, 1682-1684(2013)。
Skoglund, P.等人。从西伯利亚尼安德特人的现代污染中分离出内源性古代DNA。国家科学院学报美国111, 2229-2234(2014)。
Renaud, G., Slon, V., Duggan, A. T. & Kelso, J. Schmutzi:污染估计和内源性线粒体共识呼吁古代DNA。基因组医学杂志。16, 224(2015)。
Korneliussen, T. S., Albrechtsen, A. & Nielsen, R. ANGSD:下一代测序数据分析。BMC生物信息学15, 356(2014)。
黄,Y. & Ringbauer, H. hapCon:通过从参考单倍型复制估计古代基因组的污染。生物信息学38, 3768-3777(2022)。
Nakatsuka, N.等。污染:利用断裂连锁不平衡估计古代核DNA污染。基因组医学杂志。21, 199(2020)。
李,H.等。序列对齐/映射格式和SAMtools。生物信息学25, 2078-2079(2009)。
卡塔拉诺,G.等。地中海中部旧石器时代晚期的狩猎采集者:来自晚期Epigravettian埋葬Oriente C (Favignana,西西里岛)的新考古和遗传数据。皮疹。Int。537, 24-32(2020)。
詹森,t.z.等人。来自咀嚼桦树沥青的5700年前的人类基因组和口腔微生物组。Commun Nat。10, 5520-10(2019)。
Key, F. M.等。人类适应的出现沙门氏菌血清与新石器化过程有关。Nat,生态。另一个星球。4, 324-333(2020)。
Rivollat, M.等。来自法国的古代全基因组DNA突出了中石器时代狩猎采集者和新石器时代农民之间相互作用的复杂性。科学。睡觉。6, eaaz5344(2020)。
Svensson, E.等人。欧洲冰期前pettera Muierii头骨基因组表现出高多样性和低突变负荷。咕咕叫。医学杂志。31, 2973 - 2983。e9(2021)。
安东尼奥,M. L.等。古罗马:欧洲和地中海基因的十字路口。科学366, 708-714(2019)。
布莱斯,S.等。古代基因组显示了新石器时代早期英国的人口置换。Nat,生态。另一个星球。3., 765-771(2019)。
布鲁内尔等人。来自当今法国的古代基因组揭示了其7000年的人口历史。国家科学院学报美国117, 12791-12798(2020)。
卡西迪,L. M.等。新石器时代社会中的王朝精英。自然582, 384-388(2020)。
González-Fortes, G.等。多瑙河下游地区新石器时代农民和中石器时代狩猎采集者多代混合的古基因组学证据。咕咕叫。医学杂志。27, 1801 - 1810。e10汽油(2017)。
琼斯,e.r.等人。旧石器时代晚期的基因组揭示了现代欧亚人的深层根源。Commun Nat。6, 8912(2015)。
琼斯,e.r.等人。波罗的海的新石器时代的过渡并不是由早期欧洲农民的混合推动的。咕咕叫。医学杂志。27, 576-582(2017)。
拉扎里迪斯等人。古人类基因组显示,今天的欧洲人有三个祖先种群。自然513, 409-413(2014)。
拉扎里迪斯等人。对古代近东农业起源的基因组研究。自然536, 419-424(2016)。
李普森等人。平行古基因组横断面揭示了早期欧洲农民复杂的遗传历史。自然551, 368-372(2017)。
van de Loosdrecht等人。更新世北非人的基因组连接了近东和撒哈拉以南非洲人的种群。科学360, 548-552(2018)。
奥拉尔德,等人。伊比利亚半岛过去8000年的基因组史。科学363, 1230-1234(2019)。
Saag, L.等人。爱沙尼亚的大规模农业是从草原上的性别歧视移民开始的。咕咕叫。医学杂志。27, 2185 - 2193。e6(2017)。
西科拉等人。西伯利亚东北部自更新世以来的人口历史。自然570, 182-188(2019)。
Skoglund, P.等人。石器时代斯堪的纳维亚的采集者和农民的基因组多样性和遗传变异有所不同。科学344, 747-750(2014)。
Yang, m.a.等人的来自亚洲的4万岁的个体为欧亚大陆的早期人口结构提供了见解。咕咕叫。医学杂志。27, 3202 - 3208。e9(2017)。
Weissensteiner, H.等人。HaploGrep 2:高通量测序时代的线粒体单倍群分类。核酸测定。44, w58-w63(2016)。
Edgar, r.c. MUSCLE:多序列比对,高精度,高通量。核酸测定。32, 1792-1797(2004)。
Kumar, S., Stecher, G., Li, M., Knyaz, C. & Tamura, K. MEGA X:跨计算平台的分子进化遗传学分析。摩尔。杂志。另一个星球。35, 1547-1549(2018)。
Rohrlach, a.b.等人。使用y染色体捕获富集来解析单倍群H2显示了新石器时代向西欧扩张的两条路径的新证据。科学。代表。11, 15005(2021)。
martininiano, R., De Sanctis, B., Hallast, P. & Durbin, R.将古代DNA序列放入参考系统发育中。摩尔。杂志。另一个星球。39, msac017(2022)。
卡门,M.等人。最近Y染色体多样性的瓶颈与全球文化的变化相吻合。基因组Res。25, 459-466(2015)。
Ringbauer, H., Novembre, J. & Steinrücken, M.远古DNA纯合子性揭示的时间亲缘关系。Commun Nat。12, 5425(2021)。
帕特森,N.普赖斯,A. L. &赖希,D.人口结构和特征分析。公共科学图书馆麝猫。2, e190(2006)。
帕特森等人。人类历史上的古老混合物。遗传学192, 1065-1093(2012)。
帕拉迪斯,克劳德,J. &斯特里默,K. APE: R语言的系统发育和进化分析。生物信息学20., 289-290(2004)。
琴塔拉帕提,M. Patterson, N. & Moorjani, P.欧洲全新世主要人类混杂事件的时空格局。eLife11, e77625(2022)。
确认
作者感谢G. Marciani和O. Jöris对考古学的评论;C. Jeong, M. Spyrou和K. Prüfer对遗传学的评论;M. O 'Reilly对Fig的图形支持。5和扩展数据图。9;MPI-SHH考古遗传学部门的整个IT和实验室团队提供技术援助;M. Meyer和S. Nagel为单链库准备提供支持;K. Post, P. van Es, J. Glimmerveen, M. Medendorp, M. Sier, S. Dikstra, M. Dikstra, R. van Eerden, D. Duineveld和A. Hoekman提供来自北海的人类标本(荷兰);m.d. Garralda和A. Estalrrich提供了来自La Riera的人类标本(西班牙);J. Górski和M. zajjcc提供了来自Maszycka洞穴的人类标本;C. Di Patti提供来自圣特奥多罗2号(意大利)的人体标本;P. blaaviviius提供了获取Donkalnis人类遗骸和新的放射性碳年代的途径;意大利文化部和维罗纳省、罗维戈省和维琴察省的考古协会(Soprintendenza Archeologia Belle Arti e Paesaggio)批准了对塔利连特2号人类遗骸的访问;对Riparo Tagliente遗址(意大利)进行调查的F. Fontana; the Friuli Venezia Giulia Superintendency for providing access to the human tooth Pradis 1; and the Soprintendenza Archeologia Belle Arti e Paesaggio for the Provinces of Barletta-Andria-Trani and Foggia for providing access to the Paglicci human remains. This project has received funding by the European Research Council under the European Union’s Horizon 2020 research and innovation programme under grant agreements no. 803147-RESOLUTION (to S.T.), no. 771234-PALEoRIDER (to W.H.), no. 864358 (to K.M.), no. 724703 and no. 101019659 (to K.H.). K.H. is also supported by the Deutsche Forschungsgemeinschaft (DFG FOR 2237). E.A. has received funding from the Van de Kamp fonds. PACEA co-authors of this research benefited from the scientific framework of the University of Bordeaux’s IdEx Investments for the Future programme/GPR Human Past. A.G.-O. is supported by a Ramón y Cajal fellowship (RYC-2017-22558). L. Sineo, M.L. and D.C. have received funding from the Italian Ministry of University and Research (MUR) PRIN 2017 grants 20177PJ9XF and 20174BTC4R_002. H. Rougier received support from the College of Social and Behavioral Sciences of CSUN and the CSUN Competition for RSCA Awards. C.L.S. and T. Saupe received support from the European Union through the European Regional Development Fund (project no. 2014-2020.4.01.16-0030) and C.L.S. received support from the Estonian Research Council grant PUT (PRG243). S. Shnaider received support from the Russian Science Foundation (no. 19-78-10053).
资金
马克斯·普朗克学会提供的开放获取资金。
作者信息
作者及隶属关系
贡献
c.p ashh, h.y., a.g., A.B.R, W.H.和J. Krause进行或监督基因分析。H. Rougier和I.C.进行了太平间实践分析。Y.H.和H. Ringbauer发明了污染估算方法。K.N V.V.-M。,R.R., T. Ferraz, R.T., D.G.D., M.L., A. Modi, S. Vai, T. Saupe, C.L.S., G. Catalano, L. Pagani, D.C. and E.A. performed or supervised laboratory work. H. Rougier, I.C., L.K., A. Morala, M. Rué, S.M., L.C., J.-B.C., E. Bocaege, S. Ricci, F.B., P. Bayle, B.M., F.L.B.-R., J.-G.B., G.O., E. Bortolini, O.B.-L., G.D., M.O., A.Z., V.S., E.S., L. Sineo, J.v.d.P., L. Pecqueur, G.M., G.G., J.-M.L., C.B.G., A.G.-O., M.P.-B., D.B., M.L.L., P. Storm, C.H., J. Kabaciński, T. Filimonova, S. Shnaider, N.B., B.G.-R., M.R.G.M., A.B.M.-A., B.L., C.A.-L., A.R., C. Polet, I.J., N. Cauwe, J. Soler, N. Coromina, I.R., R.C., G. Clark, L.G.S., M.-A.J., S. Rehart, D.T., S.B., M. Romandini, L.A., H. Bocherens, C.W., S. Villotte, J.F.-L.d.P., M.G.-P., M.A.E.-B., P. Bodu, L. Smits, B.S., R.J., J. Kozakaitė, C.C., H. Benthien, K.W., R.W.S., S.C.F., T. Schüler, C.T., D.G., F.L., A. Kotula, H.P., F.S., J. Svoboda, S. Sázelová, A.C., A. Khokhlov, N.J.C., F.V., K.H., P. Semal, B.J., A. Suvorov, R. Schulting, V.M., K.M., A.B. T.T., D.C. and E.A. assembled archaeological material, descriptions and interpretations. S.T. and H.F. produced a subset of the radiocarbon dates. A. Stoessel performed µCT scanning and advised on sampling strategy. C. Posth and H.Y. wrote the manuscript with input from T.T., E.A., W.H., J. Krause and all other co-authors.
相应的作者
道德声明
相互竞争的利益
作者声明没有利益竞争。
同行评审
同行评审信息
自然感谢Marta Mirazón Lahr, Ludovic Orlando和其他匿名审稿人对这项工作的同行评审所做的贡献。
额外的信息
出版商的注意施普林格自然对出版的地图和机构从属关系中的管辖权主张保持中立。
扩展的数据图形和表格
图1 MtDNA和y染色体的系统发育。
粗体字母指(一个) mtDNA和(b) Ychr单倍群,其方框的颜色根据扩展数据图中的图例。2.斜体中的标签表示在本研究中没有新数据的先前发表的个体。
图2不同狩猎-采集群体mtDNA和y染色体单倍群的分布。
每个彩色条的长度表示携带相应单倍群的个体的比例(每个面板右侧的图例)。每一组的人数写在每个柱状图的右边。我们只绘制了有两个以上个体的组,因此,来自GoyetQ116-1聚类的个体被包括在Fournol聚类中。
扩展数据图3F4-比较lgm前欧洲狩猎采集者与Goyet Q116-1和Sunghir的亲和力的统计数据。
颜色代表被测人群的分组,圆点代表f4-值和暗、光误差条分别为1*SE和3*SE估计的5 cM块折刀。从该图可以看出,西欧的gravettian类群(Fournol类群)与Goyet Q116-1亲缘关系密切,中欧和南欧的gravettian类群(vstonice类群)与Kostenki类群的代表Sunghir类群亲缘关系密切。详细信息在数据S中提供2.B.
扩展数据图4 34 ka前狩猎-采集血统的混合图建模。
在这个混合图中,与Zlatý k实验者有关的谱系分裂比Bacho Kiro IUP组更基本,后者贡献了tiyuan, Ust 'Ishim和Goyet Q116-1(用红线表示),但没有贡献给Sunghir组。
图5欧亚狩猎采集者中检测到的ROH片段摘要。
我们将长度超过4厘米的ROH的总量可视化(一个) lgm前的个体,(b)与Epigravettian和magdalenian相关的个体,(c)携带较高比例的sidelkino相关血统的个体,以及(d)具有较高比例的奥伯卡塞尔相关血统的个体。颜色图例载于(e).每个横条代表一个人,ROH分为四个长度类别(按颜色分组)。卢短(4-8厘米,蓝色)的推断模式是普遍的,这与大多数后来的农民人群形成鲜明对比,大多数人没有任何短的卢(见101),并证明欧亚西部狩猎-采集群体的有效人口规模较小。在每个面板中绘制50 cM的总ROH虚线,以帮助在不同y轴尺度的面板之间进行比较。分组和ROH段的详细信息在数据S中提供3.B.
扩展数据图6西欧亚主成分分析显示lgm后狩猎-采集者的遗传定位。
人类起源数据集上的现代个体(灰点)基因分型被用于定义主成分分析变异,古代基因组(彩色符号)被投射到主成分分析变异上。在人类起源数据集中,新报告的超过15,000个snp的个体以黑色轮廓和填充符号显示,如右侧的图例所示,而代表性的古代基因组则以轮廓符号显示,如PCA底部的图例所示。
扩展数据图8 14 ka后狩猎-采集者Sidelkino和Oberkassel血统比例的变化。
双变量图显示了Oberkassel和Sidelkino祖先在两个欧洲地区(经度在30度以下和30度以上)随时间的扩展。x轴表示每个测试个体/群体的平均年龄,y轴表示Sidelkino血统相对于每个群体中狩猎-采集血统总数(Oberkassel + Sidelkino)的比例。三个正方形突出了与梳状陶瓷文化(CCC)相关的波罗的海汞群,与更古老的波罗的海汞群相比,显示出sidelkino相关祖先的显著增加。
扩展数据图9描述40 ka后欧洲狩猎采集者主要遗传转化的图形摘要。
此图显示(一个)狩猎-采集遗传祖先之间的分布和相互作用(b)欧洲旧石器时代晚期(绿色块)的主要时间文化细分的简化示意图(灰色块),然后是分组的中石器时代到新石器时代块(灰色块)。x轴表示图中划分的地理区域。1, y轴表示距今一千年的时间(kBP)。在面板一个,不同祖先之间的遗传亲缘关系用粗线或深线表示,杂交用箭头表示。在面板b在美国,颜色代码并不意味着考古学上的相似性。
权利和权限
开放获取本文遵循知识共享署名4.0国际许可协议(Creative Commons Attribution 4.0 International License),允许以任何媒介或格式使用、分享、改编、分发和复制,只要您对原作者和来源给予适当的署名,提供知识共享许可协议的链接,并注明是否有更改。本文中的图像或其他第三方材料包含在文章的创作共用许可中,除非在材料的信用额度中另有说明。如果内容未包含在文章的创作共用许可协议中,并且您的预期使用不被法定法规所允许或超出了允许的使用范围,您将需要直接获得版权所有者的许可。要查看此许可证的副本,请访问http://creativecommons.org/licenses/by/4.0/.
关于本文

引用本文
波施,C.,于,H.,加利奇,A.。et al。旧石器时代晚期至新石器时代欧洲狩猎采集者的古基因组学。自然615, 117-126(2023)。https://doi.org/10.1038/s41586-023-05726-0
收到了:
接受:
发表:
发行日期:
DOI:https://doi.org/10.1038/s41586-023-05726-0
这篇文章被引用
古代基因组显示了人类是如何逃脱欧洲的严寒的
自然(2023)
一名2.3万年前的南伊比利亚人将末次冰期前后生活在西欧的人类群体联系在一起
自然生态与进化(2023)
