








摘要
虽然气候变化严重影响一些水生生态系统,但它也可能与人为因素相互作用,加剧其影响。在干旱气候下,大坝会在干旱时期引起水文干旱,因为大坝排水量大幅减少。然而,尽管气候变化预计会增加干旱的持续时间和强度,但这些严重水文干旱对蝗虫动物的影响却鲜有文献记载。我们在这里记录了2011-2018年期间,阿尔及利亚东北部一个高度改良的流域的大坝排水如何受到气候变化的影响,以及一种地方性濒危的豆豆蝇,Calopteryx exulSelys, 1853,对水文干旱事件的反应。分析基于以下数据的汇编:气候(温度、降水和干旱指数)、大坝管理(水深和排水量和频率)、调查数据c . exul发生和捕获-标记-再捕获(CMR)。研究期间的特点是2014年至2017年期间发生了严重干旱,导致大坝水深降低,排入河流的水量减少,以及相关的水化学变化,特别是在2017年和2018年。这些事件可能导致了一些种群的灭绝c . exul在塞布斯河(阿尔及利亚)。CMR调查显示,该物种对水深波动敏感,避免低水位和高水位(干旱和洪水)。这项研究表明,气候变化与人类对水的需求相互作用,并影响河流流动状况、水化学和水生动物。由于未来干旱事件可能会增加,目前的研究强调了迫切需要新的lottic栖息地管理计划,以维持这一物种和可能的其他物种。
简介
地中海地区是生物多样性的热点地区,其特点是温暖、干燥和快速波动的气候1,2.这一地区尤其受到热浪和干旱期的强度和长度迅速增加的威胁3..北非尤其受到干旱的严重影响4,威胁到湿地完整性和淡水生物多样性的持续存在5.尽管当地的生物多样性已经适应了干旱和热浪6然而,目前尚不清楚物种的可塑性和适应机制是否足以有效地应对环境的快速波动7,8.此外,依靠永久水生存的物种,如许多枇枇花物种,更有可能逃离栖息地丧失的危险9,10因此需要特别注意。
自由流动的河流以物理化学条件的梯度为特征,这种梯度支配着从上游到下游的脊椎动物和无脊椎动物的生物梯度11.然而,世界上大多数河流都受到大坝的干扰,这对水文造成了影响12、泥沙负荷13、鱼类和无脊椎动物迁徙14即种群的遗传多样性15,以及其他生态过程16.河流筑坝和干旱的共同影响,干旱在炎热的气候中是常见的17,可能会加剧气候变化对生态的影响,并可能导致水生动物的灭绝18.例如,在气候温暖干燥的葡萄牙西南部,Alcántara大坝的建设导致下游塔古斯盆地干旱的持续时间和程度增加17可能会对河岸木本植被造成影响19鱼类群落20..
在北非,既有临时河流,也有永久河流。在干旱年份,永久河流的水位和流量会随着河流栖息地物理和化学方面的变化而大幅下降。考虑到lotic大型无脊椎动物群落高度依赖河流系统21,22,23在美国,与干旱相关的生态变化可以强烈地影响物种的组成和丰度24.然而,河流大型无脊椎动物也受到其他人为因素的影响25,26特别是土地利用、污染和灌溉用水,它们独立或共同改变了水道的结构完整性,以及生态系统的物理化学特征27,28,29,30.,31.了解干旱和其他人为因素对水生大型无脊椎动物种群的影响,对于预测湿地环境中生物群落的变化至关重要。这些信息尤其重要,因为极端干旱事件的频率和严重程度都在增加32以及由于人口增长的人为因素,都是预期的。
在阿尔及利亚,过去十年在塞布流域对蜻蜓区系进行了广泛调查33,34,35,显示出相当大的多样性和地方性。共有45个物种被记录下来,包括罕见的北非特有物种Calopteryx exulSelys 185335.虽然该物种曾经广泛分布在北非,但由于人类活动,其分布已经经历了实质性的收缩36.目前该病仅限于摩洛哥北部、阿尔及利亚北部和突尼斯西北部的少数孤立人群。这种豆娘自1910年以来就没有在阿尔及利亚被记录过37在2007年之后才发现了不同的种群34.最近,在阿尔及利亚中北部发现了一个新的种群38.尽管该物种目前在国际自然保护联盟红色名录上被列为濒危物种36在美国,没有采取实质性的保护措施来管理其栖息地或维持当地种群,在过去十年中,塞布斯流域的几个亚种群已经消失34.虽然人为生境退化可能是影响种群持久性的一个关键因素,但严重的干旱也可能极大地影响蝗虫动物。在这里,我们评估了极端干旱(2014-2017年)如何影响塞布斯河的物种分布。
因为干旱会影响水位、流速,以及豆娘产卵的寄主植物。植物繁殖季节的长时间干旱c . exul可能会导致不同地点的一系列灭绝。在这里,我们评估了水文动力学对c . exul结合大坝水管理的历史数据、物种分布的调查数据、飞行期间成虫的每日丰度估算和成虫的捕获-标记-再捕获。具体来说,我们利用历史气候数据和干旱指数调查了干旱强度的潜在变化,估计了大坝水位和流量的时间模式,并跟踪了阿尔及利亚东北部Seybouse河的种群持续和灭绝(图2)。1).我们还估算了不同水位条件下雄性和雌性的成虫丰度和重获率。该研究使用不同方面的数据来阐明气候变化和大坝水管理之间的潜在相互作用c . exul蝗虫生境的种群持久性。

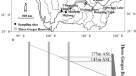
一些环境特征和地理位置的历史记载Calopteryx exul在阿尔及利亚东北部的塞布斯流域。(一个)年平均气温(B)年降水量,及(C人类足迹指数(HFI)。温度和降水数据表示30年(1970-2000年)的平均值,来自WorldClim v277.HFI范围从0(低人为影响)到100(极高人为影响)79.红圈是历史记录c . exul.倒三角表示布哈姆丹大坝。这些地图是使用R 4.0.2软件生成的(https://www.r-project.org/)和包装光栅(https://rspatial.org/raster).
结果
气候变化
1980-2018年,Seybouse流域年平均气温(Tm)呈显著上升趋势,时间斜率为0.03°C /年−1(LM: t = 5.64, P < 0.00012= 0.46)。2a). 2009-2018年间,Tm年均增加0.04°C−1,但无统计学意义(t = 1.58, P = 0.15, R2= 0.23)。对同一十年的年极端温度(最小[Tmin]和最大[Tmax])的分析表明,Tmin没有显著变化(斜率= 0.004°C年)−1, t = 0.17, P = 0.86, R2~ 0.00), Tmax呈边际增加(斜率= 0.07°C年)−1, t = 2.11, P = 0.06, R2= 0.28)。1980-2018年,Seybouse流域年降水量波动较大,但总体上无显著性变化(t =−0.51,P = 0.61, R2~ 0.00)(图;2b).然而,在2009年至2018年期间,年降水量呈−10.5 mm的时间下降−1(t =−2.75,P = 0.02, R2= 0.49)。数字2c为1980-2018年的一个月SPEI值。总体而言,近年来(2009-2018年)SPEI呈下降趋势。SPEI的负值大于负值,表明干旱期比湿润期更频繁。2009-2013年只有7个值低于−1.5阈值(表示严重干旱)(平均值为−1.83±0.20,范围为−2.08至−1.61,N = 7),其中10个值在2014-2018年(平均值为−1.88±0.23,范围为−2.22至−1.58,N = 10)。

阿尔及利亚东北部塞布斯流域1980-2018年气候条件的变化。(一个)年平均温度(实线)、最低温度(下虚线)和最高温度(上虚线)的时间格局。过去十年用红色标出(2009-2018年)。蓝线是线性回归,灰色带是标准误差。(b)年降水量的时间格局。过去十年用红色标出(2009-2018年)。蓝线为黄土回归(灰色带为标准误差),表明过去几年的下降。(c)研究区标准化降水蒸散指数(SPEI)月值。正值(湿润期)用蓝色表示,负值(干燥期)用红色表示。虚线水平线设置为−1.5,表示严重干旱。虚线表示上一个十年(2009年)的开始。黑线为黄土回归,表明在过去十年(干旱时期)SPEI下降。
大坝水管理
2011 - 2018年大坝最高水位呈时间下降趋势(图2)。3.(LM:二次效应,t =−4.92,P < 0.0001;表格S1).快速下降始于2016年,从2016年3月(360.6 m)到2018年6月(325.1 m),水深下降了9.8%。尽管近年来灌溉用水量与前几十年相比有所下降(LM:二次效应,t =−4.02,P = 0.0004)(图5)。3.b)人类消耗流量持续增加(LM: slope = 0.37 hm)3.一年−1, t = 7.02, P = 0.0001),尽管在过去十年中增长速度较慢(图。3.c)。

Bouhamdane大坝(Guelma,阿尔及利亚)水管理的时间模式。(一个) 2011-2018年最高水位逐月变化。蓝线是二次回归,灰带是标准误差。(b) 1990-2018年灌溉排水量估算。蓝线为黄土回归,灰色带为标准误差。过去十年用红色标出(2009-2018年)。(c) 1990-2018年估计供人食用的排放量。蓝线是线性回归,灰带是标准误差。过去十年用红色标出(2009-2018年)。
灌溉用水量是春、夏、秋三季的周期性事件。S1);繁殖期:该物种出现和繁殖的时期2011-2018年平均释放量最大的是7月(6.83±2.87 hm)3.)和8月(5.89±2.82 hm .3.),中间量在春末释放(5月:3.54±3.64 hm3.)、初夏(6月:4.65±3.23 hm3.),秋季(9月:4.46±2.83 hm3.10月:2.83±2.32 hm3.).2011年至2016年,月释放峰值为7.2 hm3.2010年7月为9.1 hm3.2016年6月有小流量,2017年5月有小流量(0.26 hm3.)和6月(1.74 hm .3.), 2018年没有排放,导致塞布斯河长期干旱。
化学参数
为了确定处理前原水的物理化学特征在干旱期间是否具有时间模式,在2012-2018年期间评估了16个河流(流入博哈姆丹大坝)的环境变量(图2)。S2).三个参数(TAC, HCO3.−, PO43−)显著增加(P < 0.05)。四(TH, Ca2 +、铁2 +,和NH4+(P = 0.05-0.07),而其他变量没有表现出显著的模式,尽管大多数不显著的增加(如电导率,Mg2 +, Cl−, RS, P = 0.12-0.16)可能是由于样本量低(7年数据)。
豆娘分布响应
的最大数量的亚种群c . exul(N = 8;表格S2,无花果。S3a, b).在这些种群中,2011-2016年有3个是永久亚种群,而4个是短暂亚种群(消失后又重新出现)。2018年,该河流的亚种群数量下降到一个过渡亚种群,尽管该亚种群自2012年以来已经灭绝(图2)。S3c).所有三个经常发生繁殖的永久亚种群在2018年都消失了(图2)。4a、b)。因此,在2011-2018年期间,每年损失0.85个亚种群。

繁殖的Calopteryx exul在2011年的塞布斯河(一个)以及2018年长江水位的大幅下降。请注意,长叶池塘草(Potamogeton nodosus),这是该物种的首选产卵地点,它们通常漂浮在水面上,由于抽水而干燥(来源:Rassim Khelifa)。
综合整个流域的所有地点可以证明这一点c . exul2公里半径六边形内的平均人类足迹指数(HFI:一个在0到100之间变化的指标,表示人类影响)为55.5±14.9,最小值为23.4,最大值为75.6。该值对应于流域主要水道中发生的HFI的83%,平均HFI为38.2±17.4(21-93范围)。
成虫丰度和重捕率
自水位上升的繁殖场所c . exul在一个季节内有规律地波动(图;5),主要是由于大坝排水和抽水灌溉,我们研究了水位波动对物种丰度和检测(重新捕获)的影响。根据El Fedjoudj P 2011年繁殖季节的日丰度时间模式,得出成年个体(全部且仅为成熟个体)的数量。c . exul随水深的变化呈二次型(表S3).成人人数最多c . exul在水深中等(0.2 ~ 0.7 m)的日子里记录了个体数量,而在水深较低(< 20 cm)的日子里记录的个体数量相对较少,在洪水(> 1 m)期间记录的个体数量最少(图2)。6a).水位的大幅降低(干旱)或增加(洪水)导致首选繁殖基质的消失。干旱和随后的水位下降导致河岸植被远离水面,而洪水又导致产卵寄主植物淹没。即使在干旱期间记录了一些个体,它们也没有表现出繁殖行为。

河流水位的可变性岌岌可危Calopteryx exul再版,阿尔及利亚东北部的塞布斯河。(一个)小溪快干了,(b)中间水位(豆娘最喜欢的水位),以及(c)泛滥的小溪。请注意,流的三种状态是在一周内记录的(来源:Rassim Khelifa)。

溪流水深对成虫数目的影响(一个)和标记雄性(蓝色)和雌性成年(红色)的重新捕获概率Calopteryx exul(b).(一个)拟合的直线是所有个体(蓝色)和成熟个体(红色)的负二项回归。(b)拟合线是包含恒定存活的cormark - jolly - seber捕获-标记-再捕获模型的预测。
我们的分析包括313只有标记的成年人,其中56.8%至少被重新捕获过一次。重获的平均±SD数为2.78±2.46,中位数为2,范围为1-19。S4).捕获-标记-再捕获数据表明,再捕获概率的最佳模型是水深和性别的二次效应,在非常低或非常高水位的日子里记录的再捕获概率最低,而在中等水位的日子里记录的概率最高(图2)。6b;表格S4).鉴于我们的采样范围很广,男性和女性都没有出现,这表明由于水文变化,可能会扩散出采样区域。男性比女性有更大的重新捕获概率。
讨论
我们的研究表明,气候变化可能与大坝水管理产生相互作用,加剧干旱强度,并最终影响水生生物。具体而言,结果表明,长期严重干旱改变了水的化学性质,包括污染物水平的吸收,并减少了大坝排入河流的水量,但仍保持了人类消耗的水量。这极大地影响了河流的水位和流量,从而影响了濒临灭绝的北非特有豆娘的分布,c . exul.我们强调,行动计划必须考虑到气候变化情景、大坝水管理计划、湿地测深和水文、灌溉需求和人为干扰39,40确定未来气候变化对淡水生态系统的影响,并有效管理生物多样性。
气候变化与大坝水资源管理
北非是世界上最干燥和最温暖的地区之一,也经历了严重的气候变化4.结果表明,2009-2018年期间,热力和水文条件波动较大,在研究期间的后期表现出更温暖的条件和更严重的干旱。在这一地区,大多数淡水生态系统都存在于沿海地区,那里的生物群落被局限在撒哈拉沙漠的南部,地中海的北部,限制了物种潜在避难所的范围。这意味着应特别注意气候变化对淡水群落的影响41,42哪些物种的生理、行为和分布对环境变化特别敏感41,42.温度升高,降水减少,水的物理化学性质发生变化43可能会引起生理压力44减少了物理空间,同时也增加了竞争和捕食的脆弱性45,栖息地连通性减弱46在极端条件下会导致干旱和局部灭绝47.在这里,我们展示了人为控制河流流量和气候变化共同作用的后果,导致干旱的程度和持续时间增加,预计由于气候变化,未来几年干旱的频率将增加17.
结果表明,2017年流域水量下降,2018年极端干旱事件发生后完全停止,导致河流流量(河宽和水深)大幅减少。停止放水的决定是由于大坝供人类使用的水量大幅下降。这条河被水泵进一步用于农业,导致河流的一些部分完全干旱。水量的减少以及农业径流、污水和工业排放的持续输入降低了水生动物、作物和牲畜的水质48.由于人口增长和灌溉,预计用水量会增加,这可能会加剧北非的水文干旱49.未来的模型应包括所有这些组成部分,以预测未来气候对当地蝗群的影响50.
水的物理化学性质
水化学的时间格局表明,在干旱事件期间,包括污染指标在内的许多参数都有所增加。磷酸盐大幅增加到促进富营养化的水平(> 0.1 mg/L)和其他不利的环境影响51.氨,高浓度时对水生动物有毒52,近年来也有所增加。水化学的这些变化可能是由于干旱造成的,干旱降低了水位,增加了水中营养物质和有毒化合物的浓度。这些数据仅用于推断大坝中水化学和水质的潜在时间变化,而不是下游所遇到的水平c . exul发生。例如,2013年处理前原水中记录的磷酸盐、氨和硫酸盐水平分别比Medjez Amar(上游Seybouse河)记录的水平低60倍、50倍和3倍53,分别。这种增长是由于大量人口产生的污水、农业的集约土地使用和工业污染。因此,干旱不仅会影响水的物理性质,还会影响水的化学性质,使蝗虫栖息地的环境条件非常紧张,并使水生动物超出其生理耐受范围。
影响Calopteryx exul人口
2014-2017年的严重干旱可能直接和/或间接导致了濒危豆娘不同种群的灭绝c . exul因为它与人口的快速流失同时发生。的分布c . exul在过去的十年里,塞布斯河的淡水资源迅速减少34.根据国际自然保护联盟(IUCN)的B2和E标准,考虑到该物种的活动范围大小、栖息地破碎化的严重程度和过去十年的局部灭绝概率,目前该物种在该地区的迅速灭绝率表明,该物种应在当地被提升为极度濒危物种。目前的研究表明,在过去十年中表现出最具适应力的亚种群34在2018年被灭绝,剩下的种群定居在过去经历过灭绝的地点。需要指出的是,该物种可能在流域的其他地方找到了避难所,这值得在未来的监测中进行进一步调查。然而,该物种在塞布斯河的未来生存将受到未来干旱事件以及为满足人类需求而进行的大坝水管理的威胁。
自c . exul干旱在整个生命周期中高度依赖流动的水,干旱本身(不包括其他相互作用)可能对所有生命阶段的健康产生影响。首先,卵通常产在靠近水道岸边的浮叶中34.在干燥条件下,水位的降低会导致植物脱水,最终导致虫卵死亡。其次,通常生活在流速相对较快的水中的幼虫也受到干旱的严重影响,因为水速和物理水生空间都在下降。虽然幼虫可以在潮湿的地区找到避难所,但长期干旱可能会因捕食或干燥和其他非致命影响(能量储备耗尽和体型缩小)而导致死亡。第三,即使成虫适应了在干旱的情况下分散,并寻求更适合繁殖的地点或好客的避难地点撤退但不繁殖54在美国,干旱会对该物种的种群动态产生重大影响。成熟成虫的寿命很短(< 1个月),因此该物种处于“与时间赛跑”的状态,繁殖季节的每一天在该物种的整体种群动态中都很重要。在长期干旱期间,合适的繁殖栖息地变得稀缺甚至消失,雌性在死亡前只会产下一小部分卵,如果有的话。这会影响最初的种群大小(存活卵的数量),从而影响次年或同年第二个产卵季节的新出现种群的大小55.
不幸的是,繁殖季节c . exul与旱季和农业抽水需求的增加重叠。塞布斯河的边界是种植各种作物和水果的农田,其中一些对水的供应要求很高(如西瓜、西瓜和西红柿)。这意味着已经存在大量的灌溉用水,这降低了河流和溪流的水位。在干旱年份,河流水量减少,但抽水需求保持不变,这增加了对水文的影响,并对水生和陆地群落造成级联效应56.在一段极度干旱的时期之后,大坝水量大幅下降,威胁到水安全,大坝停止了所有的排水,农民们开始开采河流,直到它完全干涸。博哈姆丹大坝位于塞布兹河的一条支流上,而另一条主要支流(切尔夫河)也有一座大坝(Foum El-Khanga大坝),容量为1.57亿米3..这座大坝离其余的大坝15公里远c . exul在Ain Makhlouf附近的Cherf河上,由于一座桥梁的建设,它存在于一个高度退化的地点。因此,塞布斯流域所有已知的亚种群都受到大坝和干旱的双重影响。
除干旱外,大坝泄洪还改变了水流动态,推动了洪水的发展c . exul到不适宜居住的水文条件的另一个极端。洪水可能使幼虫漂流到食物供应和水质较低、死亡风险较高的不适宜的栖息地57,58.洪水经常改变河岸植被的群落结构,减少产卵地点的可用性。因为该物种对水流和岸边植被等栖息地特征具有选择性33时,由于水速超过最佳条件,雄鱼在高水位时不表现出生殖行为(领地性),雌鱼停止产卵。未来预测该物种种群动态的模型不仅应考虑到干旱,而且还应考虑到洪水事件,这些事件是影响所有生命阶段种群参数的重要因素。
Capture-mark-recapture
通过对捕获标记-再捕获数据的分析,进一步证明该物种对水文制度敏感。成虫的捕获概率与河流水深有显著的关系。在两性中,当水位非常低(干旱)或非常高(洪水)时,重新捕获率都大大下降。这表明该物种喜欢中等水深(也与中等水速有关),可能是因为它为产卵和卵和幼虫的生存提供了最佳条件59.在非最优条件下,c . exul似乎是为了寻找其他潜在的地点而分散,并逃避不利的当地环境变化,这与其他种类的Calopterygidae相似60,61,62.先前对该物种的研究表明,栖息地的改善(由于繁殖领地的可用性增加)增加了重新捕获的概率,增加了当地成年种群的规模,以及繁殖事件的数量34,63.平均重捕率的性别差异c . exul是常见的,这是由于行为差异,雄性是领土,更明显,并永久靠近水,而雌性更隐蔽,可能在产卵后离开水64.然而,两性对大坝水管理导致的河流水位变化的反应相似。目前尚不清楚在栖息地质量退化(干旱或洪水)后殖民的新斑块是否只是暂时适合繁殖,或者沉积的卵是否能存活到羽化。
管理的角度
干旱是地中海气候中的常见现象,未来干旱的频率和持续时间都将增加65.严重干旱会加剧其他干扰因素的影响,如污染和灌溉用水。必须采取有效措施,管理用于消费和灌溉的水供应,同时保护河流中的脊椎动物和无脊椎动物66.我们讨论了一套可以改善保护状况的行动c . exul并增强蝗虫生物多样性对极端天气事件的抵御能力。
建立一个人工沟渠系统,在极端干旱的情况下,既可用于灌溉,又可用于蝗虫动物的避难所,可以减轻干旱对蝗虫生态系统的影响67.在南非,一些开普特有的物种在严重干旱时占据人工水库,并将其作为避难所,直到它们的繁殖区恢复有利条件68.这种逃避行为是典型的适应干旱的地方性蝗虫,它们依赖于多年生河流和溪流来繁殖和维持存活的种群。理想情况下,在流域尺度上的人工水道网络(保护走廊)可以让物种分散、躲避干旱,并有可能找到合适的斑块来建立种群69.如果这些人造地点支持一些有吸引力的栖息地元素,如岸边的植被,以及保护产卵地点的漂浮树叶,这些人造地点的成功率可能会提高。例如,c . exul成虫可以通过提供产卵地点(漂浮的植物叶子)来吸引到特定的区域。34,63,当水位恢复正常时,它们会被移回原来的位置。
在北非,lottic栖息地比死水栖息地受到的保护关注少得多。认识到斑块生态系统的生态和社会经济重要性是在当地流域建立保护区的基本目标70.在这种情况下c . exul,鉴于近年来它的几个亚种群已经消失,有一种挽救它免于灭绝的方法是将其指定为保护区,因为那里的河岸干扰较少。这种栖息地保护也可能产生保护伞效应,使其他受威胁的地方性蜻蜓受益,例如Gomphus lucasiiSelys, 1849年,以及广泛的无脊椎动物和脊椎动物。
而c . exul可能对干旱有一定的恢复力,从河流和溪流中抽水是一种补充压力,可能会通过延长干旱期将物种推向恢复力的极限,并对水外的幼虫产生不利影响。水道改造是改变水流速度和改变水生大型无脊椎动物栖息地完整性的另一个因素71.需要严格的规定来限制从自然栖息地抽出的水量。此外,灌溉的替代水源,如适当引导大坝水的人工沟渠系统,是解决问题的关键。
鉴于大部分流域都被农业用地占用,制定新的保护和恢复措施,促进生态友好的耕作方式和更自然的景观配置,应该是战略保护计划的首要任务72.将严重依赖化肥和农药的耕作方式转变为更多的有机耕作,很可能会减少流入河流和溪流的有毒物质,改善繁殖地点的水质,改善觅食地点的栖息地质量,并改善食物质量(未受污染的猎物)。73,74.由于流域内的农业用地是高度简化的单一栽培,栖息地复杂性非常低,在原生草地旁边添加树木、灌木和混合作物等栖息地元素将改善景观复杂性,维持生物多样性,并增强生态系统的恢复力75.这种向多样化农业生态系统的转变应在提高公众和其他利益攸关方的认识以及随后的政策方向之后逐步实施。
在流域范围内开展定期长期监测方案对于评估种群趋势、物种对环境和人为扰动的敏感性以及新实施的管理计划的成功至关重要。此外,即使保护当局不会对流域保护投入太多的注意力和资源,一个精心策划的社区科学计划也可以在数据收集、公共教育和保护宣传方面发挥相当大的作用。最终,对河流生态系统以及未来气候影响的有效管理需要一个涉及生态学、气候学、水文学、社会学、经济学等多个学科科学家的多学科合作网络。不幸的是,尽管局势紧迫,但我们目前还远远没有在塞布斯河沿岸建立这样的网络。
方法
研究网站
该研究是在位于阿尔及利亚东北部的Seybouse河进行的。1).这条河流入地中海,由Bouhamdane河和Cherf河在Guelma市以西10公里处的Medjez Amar汇合而成。
Bouhamdane大坝位于阿尔及利亚Guelma以西23公里的Hammam Debagh省(36°27′40″N, 7°14′15″E)。大坝位于塞布兹河源头(36°26′35″N, 7°18′39″E)上游7公里处的博哈姆达姆河上,限制了流入塞布兹河的博哈姆达姆河的流量。大坝的表面积为1070公里2最高水位370米。它的容量为220 hm3.速度是2240米3.年代−1.它为不同省份(Guelma, Ben Djerrah, Medjez Amar, Hammam Debagh和Ain Hessainia)提供饮用水,这些省份估计需要9 hm3.一年−176.该地区的雨季(温和湿润的冬季)为10月至4月/ 5月,旱季(炎热干燥的夏季)为6月至9月,与包括蜻蜓在内的大多数水生昆虫的繁殖季节一致35.图中显示了Seybouse流域的气候和人为特征。1.
气候和大坝数据
为了表征Seybouse流域的区域气候条件,使用了从WorldClim v2上获得的2.5 min空间分辨率的年平均温度(Tm)和年降水量(P)数据(1970-2000年和Seybouse流域的平均值)77.1980-2018年历史月度天气数据78对塞布斯流域的Tm和p的时间趋势进行了计算,以确定人类对流域和c . exul全球人类足迹指数(HFI),空间分辨率为1公里2使用79.HFI是一个多元指标,包括人口密度、人类土地利用、基础设施干扰(如建筑物、夜间人工照明)和人类通道(如道路、铁路)。评估人类活动对某地的影响强度c . exul在塞布斯流域的水系统中,平均HFI在4公里处2-六边形网格计算。标准化降水蒸散发指数(SPEI)是衡量干旱严重程度(强度和持续时间)的指标,从全球SPEI数据库中获得了研究区域(36.25°N, 7.25°E)的一个月值80.使用一个月的SPEI(而不是3个月和6个月)来避免时间自相关。阈值为−1.5被用作严重干旱的指标81.为了评估大坝水位和流量的时间变化,从大坝管理人员那里获得了2011-2018年博哈姆丹大坝的月最高水位和月排水量的数据。此外,为了确定水的物理化学性质在干旱期间是否发生了变化,我们使用2012-2018年期间调查的16个参数的年平均值,即pH值、浑浊度、电导率、总碱度(TAC)、总硬度(TH)、碳酸氢盐(HCO),跟踪了处理前布汉丹大坝原水的水物理化学特征的时间趋势3.-),钙(Ca2 +)、镁(Mg2 +),氯(Cl−)、干渣(RS)、硝酸盐(NO3.−),铁(Fe2 +),硫酸盐(SO42−),磷酸(PO43−),铵(NH4+)、有机物(MO)。
抽样方法
的亚种群进行了调查c . exul在2017年和2018年的繁殖季节。用200米的横断面沿河道河岸记录成年个体(幼体、未成熟和成熟)的发生情况。此外,我们使用了早在2011年的地点的物种发生数据,当时塞布岛的亚种群数量最高34.在繁殖季节,每个亚种群至少被访问三次。在本文中,我们只评估了布哈姆丹河和塞布斯河的亚种群,因为后者对布哈姆丹大坝的水管理很敏感。
在2011年繁殖季节,在塞布兹河上游开展了广泛的捕获-标记-再捕获计划,以估计种群规模,评估出现模式,描述繁殖行为,了解物种的传播习惯35,55,82.2011年4月28日至5月29日,每日捕捉c . exul成虫是在河道中制造的(El Fejdoudj P;36°28′21″N, 7°22′15″E)在上午(09:00-12:00)横穿100 m样带。在用手网捕获豆娘后,用永久记号笔在翅膀上标记单个的字母数字代码。标记只花了几秒钟,昆虫立即被释放到相同的位置。每天进行重新捕获(重放)。同时还记录了整个样带的成虫总数(雄性和雌性)。由于该物种在形态和行为上都很明显,很有可能在该地点记录了所有的个体。由于研究地点的水深每天都在波动(图。5),在研究河道水床中心的特定位置,使用分级棒每日估算水深。水床中心水位的波动反映了河道水文的一般变化。我们利用313个标记个体的捕获历史,测试了这种水文波动与重新捕获率变化的相关性。值得注意的是,特定地点水深的变化也是水流变化的一个指标;一个重要的环境特征,它高度影响a的可能性Calopteryx物种在特定的地点繁殖59.
统计分析
所有分析均采用R 3.2.3进行83.采用多项式回归法对大坝水位的年变化进行分析。采用线性回归方法对流域年降水量、年平均气温、年最低气温、年最高气温、年降水量的变化进行了评价。采用cormark - jolly - seber (CJS)模型对捕获-标记-再捕获数据进行分析RMark包84.用函数检验了模型的假设release.gof提供评估独立性和短暂性的测试(TEST2、TEST3和TOTAL)85.所有检验均无显著性(P > 0.50),表明数据满足CJS模型的要求86.由于重新捕获概率(p)是感兴趣的变量,因此生存概率(Phi)固定为1,并测试了个体(性别)和环境协变量(水深[WD])的影响。候选模型的选择从一个简单模型开始,包括一个协变量(性别和水深)的线性影响,到一个更复杂的模型,包括水深的多项式项(WD + WD)2),以及协变量(WD + WD .)的加性效应2+性)。首先对模型进行计算,然后根据校正赤池信息准则(AICc)进行模型选择。使用负二项回归分析了水深对跨越水道的成人数量的影响。数值为平均值±标准差。
数据和材料的可用性
数据存储在GitHub存储库中:https://github.com/rassimkhelifa/Data.
参考文献
迈尔斯,N.,米特梅尔,R. A.,米特梅尔,C. G.,达·丰塞卡,G. A. &肯特,J.生物多样性保护优先热点。自然403, 853-858(2000)。
Lionello, P。et al。在地中海气候变化第4卷(编莱昂内洛,P。et al。) 1-26 (Elsevier, 2006)。
Molina, M, Sánchez, E. & Gutiérrez, C.从euro-coRDeX区域气候模式集合预测地中海上空的未来热浪。科学。代表。10, 1-10(2020)。
布奇尼亚尼,E.,麦科利亚诺,P.,帕尼茨,H.-J。用cosmom - clm在不同空间分辨率下对中东-北非地区的气候变化预测。放置爬。改变9, 66-80(2018)。
García, N., Cuttelod, A. & Malak, D. A.;北非淡水生物多样性现状与分布(IUCN, 2010)。
迪·卡斯特里,F. &穆尼,H. A。地中海型生态系统:起源与结构第7卷(施普林格科学与商业媒体,2012)。
Stoks, R., Geerts, A. N. & De Meester, L.淡水无脊椎动物对气候变化的进化和塑料响应:已实现的模式和未来潜力。另一个星球。达成。7, 42-55(2014)。
韦伯恩,G. A.,斯凯利,D. K. &沃纳,E. E.跨淡水生境梯度创造群落结构的机制。为基础。启生态。另一个星球。系统。27, 337-363(1996)。
Arribas, P。et al。静水甲虫和潮水甲虫(鞘翅目:亲水甲虫科)活动范围的差异是由其分散能力而非生态耐受性决定的。j . Biogeogr。39, 984-994(2012)。
Hof, C., Brändle, M. & Brandl, R. lentitic odonates比lotic种有更大更北的活动范围。j . Biogeogr。33, 63-70(2006)。
范诺特,r.l.,明肖尔,g.w.,康明斯,k.w.,塞德尔,j.r.和库欣,c.e.河流连续统概念。可以。j .鱼。Aquat。科学。37, 130-137(1980)。
Ibàñez, C., Prat, N. & Canicio, A.埃布罗河下游及其河口大坝产生的水文和泥沙运输变化。Regul。Rivers Res. Manag。12, 51-62(1996)。
孔道夫,G.,鲁宾,Z. &米尼尔,J.湄公河大坝:累积泥沙饥饿。水Resour。Res。50, 5158-5169(2014)。
林志刚,李志刚,李志刚。新世界水文变化对河流大型生物群的区域影响:热带-温带比较。生物科学50, 807-823(2000)。
刘,X。et al。鄱阳湖流域大坝及其环境影响对淡水贻贝种群遗传多样性和连通性的影响Freshw。医学杂志。65, 264-277(2020)。
巴巴罗萨,V。et al。当前和未来大坝对全球淡水鱼地理范围连通性的影响Proc。国家的。学会科学。美国117, 3648-3655(2020)。
López-Moreno, j。et al。大坝对跨界盆地干旱程度和持续时间的影响:塔古斯河下游,西班牙和葡萄牙。水Resour。Res。45, w02405(2009)。
McMahon, T. & Finlayson, B.干旱和抗旱:澳大利亚河流的低流量水文。Freshw。医学杂志。48, 1147-1160(2003)。
阿吉亚尔,F. C. &费雷拉,M. T.人为干扰景观:对葡萄牙塔古斯河流域河岸木本植被组成和完整性的影响。环绕。Conserv。32, 30-41(2005)。
Costa, M. J., Vasconcelos, R., Costa, J. & Cabral, H.河流流动对塔古斯河口鱼类群落的影响(葡萄牙)。Hydrobiologia587, 113-123(2007)。
生物群落有效性对南非河流中大型无脊椎动物组合的影响:对水生生物评估的影响。Freshw。医学杂志。52, 370-380(2007)。
Demars, B. O., Kemp, J. L., Friberg, N., Usseglio-Polatera, P. & Harper, D. M.河流中生物群落与无脊椎动物的联系:生物学特征,分类组成和多样性。生态。印度的。23, 301-311(2012)。
大型无脊椎动物群落从干扰中恢复的研究。环绕。等内容。14, 605-620(1990)。
干旱对河流大型无脊椎动物组合影响的平行与对比。Freshw。医学杂志。48, 1173-1185(2003)。
Desrosiers, M。et al。利用底栖大型无脊椎动物的特征评估圣劳伦斯河的人为压力。科学。总环境。649, 233-246(2019)。
杜伦斯,I. & Ormerod, S. J.气候变化对旱流大型无脊椎动物25年的影响。水珠。改变医学杂志。13, 942-957(2007)。
桑托斯,R。et al。气候变化和土地利用情景对濒危物种Margaritifera的影响科学。总环境。511, 477-488(2015)。
朱尼尔,r.f.v。et al。土地利用冲突对河流生态系统的影响土地使用政策43, 48-62(2015)。
Fonseca, A, Fernandes, L. S, fontainas -Fernandes, A, Monteiro, S. & Pacheco, F.淡水金属浓度对鱼鳃组织病理学变化严重程度的影响:统计角度。科学。总环境。599, 217-226(2017)。
费雷拉,费尔南德斯,L. S.科尔特斯,R. &帕切科,F.利用嵌套偏最小二乘回归评估人类活动对河流生态系统的影响。科学。总环境。583, 466-477(2017)。
费尔南德斯,L. S.,费尔南德斯,A.,费雷拉,A.,科尔特斯,R. &帕切科,F.葡萄牙农村和城市流域生物多样性损失的偏最小二乘路径建模分析。科学。总环境。626, 1069-1085(2018)。
政府间气候变化专门委员会。2014年气候变化:影响、适应和脆弱性:区域层面(剑桥大学出版社,2014)。
Khelifa, R.飞行时期,明显的性别比例和栖息地偏好的马格里比亚特有Calopteryx exulSelys, 1853(蜻蜓目:颧翅目)。生态讽刺剧《地球与生活》68, 37-45(2013)。
Khelifa, R. & Mellal, M. K.宿主植物修复作为改善o捐赠专家保护状况的潜在工具。昆虫Conserv。潜水员。10(2), 151 - 160。https://doi.org/10.1111/icad.12212(2017)。
Khelifa, R。et al。塞布兹河(阿尔及利亚东北部)受威胁的地中海蜻蜓的热点:IUCN的种群规模被严重低估了吗?Int。j . Odonatol。19, 1 - 11。https://doi.org/10.1080/13887890.2015.1133331(2016)。
Boudot, j。Calopteryx exul。2018年世界自然保护联盟濒危物种红色名录e.T60287A72725790。https://doi.org/10.2305/IUCN.UK.2018-2301.RLTS.T60287A72725790.en.下载于1月72725729 72722021。(2018)。
Martin R.投稿à l’étude des Neuroptères de l’africa。2Les oddonates du département de Constantine。安。Soc。Entomol。Fr。79, 95-104(1910)。
Chelli, A., Zebsa, R. & Khelifa, R.在阿尔及利亚北部中部发现了一种濒临灭绝的新种群Calopteryx exul(蜻蜓目:Calopterygidae)。不是。Odonatol。9, 150-154(2019)。
费燕,李志刚,李志刚。全球变暖对欧洲河流干旱的影响。j .地球物理学。研究大气压。114, d17116(2009)。
施耐德,C., Laizé, C.,阿克雷曼,M. &弗洛克,M.气候变化将如何改变欧洲的河流流动状况?二聚水分子。地球系统。科学。17, 325-339(2013)。
愤怒,D。et al。淡水生物多样性:重要性、威胁、现状和保护挑战。医学杂志。牧师。81, 163-182(2006)。
斯特雷耶,D. L. &达金,D.淡水生物多样性保护:近期进展和未来挑战。J. North Am。Benthol。Soc。29, 344-358(2010)。
范弗里特,M. & Zwolsman, J.夏季干旱对默兹河水质的影响。j .二聚水分子。353, 1-17(2008)。
Caruso, B.极低流量的时空模式和对新西兰奥塔哥溪流生态系统的影响。j .二聚水分子。257, 115-133(2002)。
史丹利,费希尔,s.g. &格林,n.b.溪流生态系统的扩张与收缩。生物科学47, 427-435(1997)。
Truchy,。et al。生境斑块,生态连通性和北方河流生态系统从实验干旱的不均匀恢复。水珠。改变医学杂志。26, 3455-3472(2020)。
博尔顿,a.j. &雷克,p.s.。干旱对溪流昆虫的影响及其生态后果。水生昆虫:种群的挑战81-102 (cabi, 2008)。
安德森,C. B.,刘易斯,G. P. &萨金特,K. A.废水处理废水对浓度和溶质通量的影响布什河,南卡罗来纳州,在极端干旱条件下。环绕。Geosci。11, 28-41(2004)。
Wada, Y., Van Beek, L. P., Wanders, N. & Bierkens, M. F.人类用水加剧了全球水文干旱。环绕。卷8, 034036(2013)。
Aldous, A., Fitzsimons, J., Richter, B. & Bach, L.干旱、洪水和淡水生态系统:评估气候变化的影响和制定适应策略。3月Freshw。Res。62, 223-231(2011)。
康利,d.j.。et al。控制富营养化:氮和磷。科学123, 1014-1015(2009)。
公园,T.-J。et al。用物种敏感性分布法建立保护淡水生物的氨水质标准。科学。总环境。634, 934-940(2018)。
雷根,A.,布彻拉赫姆,E.-H。,Hanane, S. & Houhamdi, M. Effects of anthropogenic activities on the quality of surface water of Seybouse River (northeast of the Algeria).阿拉伯人。j . Geosci。10, 219(2017)。
Khelifa, R。et al。一种濒危的地方性豆娘的远距离运动Calopteryx exulSelys, 1853(卡翅目:蜻蜓目)。误判率。j .生态。52, 375-377(2014)。
北非特有豆娘的部分双翅性和出现模式Calopteryx exul:对保育的影响。误判率。j .生态。55, 145-151(2017)。
亚当斯博士。et al。干旱引起的树木死亡的温度敏感性预示着在全球chang型干旱下区域死亡的增加。Proc。国家的。学会科学。美国106, 7063-7066(2009)。
斯克林杰,G. J. & Winterbourn . M. J.洪水对表层和底栖大型无脊椎动物种群在不稳定的新西兰河流的影响。Hydrobiologia171, 33-44(1989)。
吉勒,P. Sangpradub, N. & Twomey, H.特大洪水与大型无脊椎动物群落结构。Verh。Int。版本。定理。Angew。Limnol。241724-1729(1991)。
西瓦-乔西,M. T.吉本斯,D. W. & Pain, D.豆娘雌性产卵地点偏好和卵孵化成功萼祖鸟黄口瘤.Behav。生态。Sociobiol。37, 39-44(1995)。
施特默(C. Stettmer)。Calopteryxsplendens)和美丽的姑娘(c .处女座)(齿形目:齿形目)产于德国东南部的溪流中。欧元。j . Entomol。93, 579-593(1996)。
Chaput-Bardy, A., Grégoire, A., Baguette, M., Pagano, A. & Secondi, J.豆娘的条件和表型依赖扩散,Calopteryx splendens.《公共科学图书馆•综合》5, e10694(2010)。
沃德,L.和米尔,P.个人的长距离运动作为范围扩展的工具Calopteryx splendens(蜻蜓目:束翅亚目)。欧元。j . Entomol。104, 195(2007)。
Mellal, M. K., Bensouilah, M., Houhamd, M. & Khelifa, R.生殖栖息地供应促进濒危地方性豆娘的生存和繁殖Calopteryx exul.J.昆虫保护22, 563-570(2018)。
科德罗-里维拉,A. &斯托克斯,R. In蜻蜓和豆娘:生态和进化研究的模式生物(编者:Córdoba-Aguilar, A.) 7-20(牛津大学出版社,2008)。
Iglesias, A., Garrote, L., Flores, F. & Moneo, M.管理地中海缺水和气候变化风险的挑战。水Resour。等内容。21, 775-788(2007)。
巴奈特,t.p.。et al。美国西部水文的人为变化。科学319, 1080-1083(2008)。
萨姆韦,m.j.。et al。非洲南部干旱地区人工池塘对水生昆虫的价值综述。Biodivers。Conserv。29, 3131-3150(2020)。
Deacon, C, Samways, M. J. & Pryke, J. S.水生昆虫数量下降,并占据低质量的人工栖息地以在水文干旱中生存。Freshw。医学杂志。64, 1643-1654(2019)。
布里格斯,a . J., Pryke, J. S., Samways, M. J. & Conlong, D. E.农村景观马赛克中池塘景观蜻蜓之间的互补。昆虫Conserv。潜水员。12, 241-250(2019)。
综合淡水生态学与生物多样性保护。生态。印度的。11, 1507-1516(2011)。
布鲁克斯,A. J., Chessman, B. C. & Haeusler, T.大型无脊椎动物特征区分不受控制的河流受水的抽取。J. North Am。Benthol。Soc。30., 419-435(2011)。
洛杉矶,加里波第。et al。工作景观至少需要20%的原生栖息地。Conserv。列托人。https://doi.org/10.1111/conl.12773(2020)。
文森特,A. &弗勒里,P.为保护水质而发展有机农业:法国当地项目及其政策含义。土地使用政策43, 197-206(2015)。
Bengtsson, J., Ahnström, J. & Weibull, a.c.有机农业对生物多样性和丰度的影响:元分析。j:。生态。42, 261-269(2005)。
利希滕伯格,e。et al。多样化耕作系统对农田和农业景观内节肢动物多样性影响的全球综合。水珠。改变医学杂志。23, 4946-4957(2017)。
ABHCSM。A.G.I.R.E(国家资源署intégrée法国资源)(2016)。和谐的水文分析'année水文(2015-2016)哈马姆·德巴。君士坦丁瓦-塞布斯-梅勒格盆地水文局(2016)。
菲克,S. E. & Hijmans, R. J. WorldClim 2:全球陆地地区新的1公里空间分辨率气候表面。Int。j . Climatol。37, 4302-4315(2017)。
Harris, I., Jones, P. D., Osborn, T. J. & Lister, D. H.更新的月度气候观测的高分辨率网格- CRU TS3。10的数据集。Int。j . Climatol。34, 623-642(2014)。
野生动物保护学会- wcs和国际地球科学信息网络中心- ciesin -哥伦比亚大学(NASA社会经济数据和应用中心(SEDAC), 2005)。
Vicente-Serrano, s.m. & Staff。气候数据指南:标准化降水蒸散指数(SPEI)。检索从https://climatedataguide.ucar.edu/climate-data/standardized-precipitation-evapotranspiration-index-spei(2015)。
D 'Orangeville, L。et al。干旱时间和当地气候决定了东部温带森林对干旱的敏感性。水珠。改变医学杂志。24, 2339-2351(2018)。
在一种罕见的地方性豆娘中,雌性“协助”球鞋雄性欺骗支配性雄性:性冲突是最好的。生态One hundred., e02811(2019)。
R开发核心团队。R:统计计算的语言和环境(R统计计算基础,2020)。
Laake, J. RMark:用MARK分析捕获-再捕获数据的R接口,AFSC处理报告2013-01(阿拉斯加鱼。科学。分,NOAA,国家海洋渔业局,2013)。
伯纳姆,k.p.。基于释放-再捕获的鱼类生存实验设计与分析方法第5卷(美国渔业学会专著,1987年)。
阿姆斯特鲁普,S. C.麦克唐纳,T. L. &曼利,B. F.。捕获-再捕获分析手册(普林斯顿大学出版社,2010)。
确认
我们感谢两位审稿人提出的有益意见和建议。我们感谢所有在实地调查中提供帮助的同事。特别感谢John S. Richardson的有用建议。我们还要感谢博哈姆丹大坝(Hammam Debagh)的管理人员分享数据。作者声明没有利益冲突。由瑞士国家科学基金会(P2ZHP2_175028)资助。
作者信息
作者及隶属关系
贡献
概念化:上面;方法:上面;形式分析与调查:r.k., HM;数据收集:a.b., r.k., H.M.;可视化:r.k., H.M.;写作-初稿准备:R.K.;写作评论和编辑:R.K., h.m., a.b., r.a.c.和M.S.
相应的作者
道德声明
相互竞争的利益
作者声明没有利益竞争。
额外的信息
出版商的注意
施普林格自然对出版的地图和机构从属关系中的管辖权主张保持中立。
补充信息
权利和权限
开放获取本文遵循知识共享署名4.0国际许可协议,允许以任何媒介或格式使用、分享、改编、分发和复制,只要您对原作者和来源给予适当的署名,提供知识共享许可协议的链接,并注明是否有更改。本文中的图像或其他第三方材料包含在文章的创作共用许可协议中,除非在材料的信用额度中另有说明。如果材料未包含在文章的创作共用许可协议中,并且您的预期使用不被法定法规所允许或超出了允许的使用范围,您将需要直接获得版权所有者的许可。如欲查看本牌照的副本,请浏览http://creativecommons.org/licenses/by/4.0/.
关于本文

引用本文
哈利法,R.,马赫朱布,H.,巴卢杰,A.。et al。气候变化和人类用水需求对豆娘高度威胁的影响。Sci代表11, 7725(2021)。https://doi.org/10.1038/s41598-021-86383-z
收到了:
接受:
发表:
DOI:https://doi.org/10.1038/s41598-021-86383-z
