








文摘
Irano-Turanian和地中海地区植物区系之间的关系已经从旧。然而,只有少数生物地理学分析基于分子数据评估草原植物在地中海盆地的历史。我们的研究旨在有助于更好地理解迁徙和重建生物地理学的多元化进程Rhaponticoides(Cardueae),分布在地中海和Irano-Turanian地区。我们生成的核和质体序列,分析了贝叶斯推理。我们使用产生的发展史为约会属和检查传播途径的多样化。两个演化支被恢复,Irano-Turanian进化枝和地中海进化枝。属的起源是放置在中间的安纳托利亚高原第三纪中新世。属经历了几个多元化发展和扩张相关Messinian盐度危机和上新世和第四纪环境变化。Rhaponticoides迁移后两种路线反映了两个灵魂的属:Irano-Turanian地中海类群征服了欧亚大陆草原而物种迁移通过地中海东部和中部和北部非洲,所到之处皆物种;这两种方式在伊比利亚半岛结束。我们的研究也证实,更多的工作是需要解开的系统发育关系Rhaponticoides。
介绍
地中海盆地继电器的非凡生物多样性的环境异质性,戏剧性的环境波动,millenial人类影响和温带和亚热带地区之间的地理位置,使其张力区1,2,3。然而,当前该地区的植物区系丰富也由过去的气候和地质事件4,5。推理从palaeobotanical仍然和分子数据显示,移民海浪和物种形成和消亡过程是由四个主要事件:热带气候的冷却和aridification新第三纪期间;Messinian盐度危机;地中海气候的出现;和第四纪的arid-cold波动2,3,6。因此,地中海盆地主机类群比例明显来自不同生物地理的区域,以及大量的流行源于extra-Mediterranean血统2,7,8。
大草原植物是extra-Mediterranean地中海盆地内元素获得特殊相关性9,10贡献显著地中海植物区系的丰富性2,9。它由类群适应大陆气候昼夜之间剧烈的温度波动和不同季节(winter-summer)和高降雨量季节性,通常从高原和山脉在开放的栖息地9,10,11。草原植物展品高多样性在亚洲中部和西部,阿拉伯半岛和非洲北部12,13。在这个广阔的范围,Irano-Turanian地区(中部和西部亚洲)是主要的核心在全北区的王国已经出口许多草原分类群在多个波相邻地区在新生代14,15。Irano-Turanian地区和地中海盆地众多旱生植物,达到其在伊比利亚半岛的最极端,摩洛哥北部和南部Macaronesia16。
有趣的存在草原植物在地中海盆地地解释了Messinian模型14,这主要是基于Messinian盐度危机发生在Miocene-Pliocene边界17。古证据支持部分干燥的地中海和不可告人的恢复14,17。这两个事件将会促进东西方迁移亚洲草原植物和随后的分散分布的几个类群在Mediterranenan盆地和亚洲14,18。这个合成的地理模式被公认为“Kiermack分离”19。一些研究利用花粉化石和分子数据支持Messinian盐度危机之间的联系(7 - 5 Ma)和西方之间的合成分离模式和东地中海范围,亚洲和西方7,15,20.。相比之下,其他研究显示,最近整个第四纪气候和海平面振荡可能也促进草原植物的迁移和现存的碎片在地中海盆地18,21。地中海盆地不是被冰覆盖和冻土大部分欧洲中部和北部的冰川时期,但它确实当前草原的气候类似的经验22。同样,在这些寒冷期海平面下降了近130,支持土地在地中海的出现23,24。因此,适宜的气候和土地桥梁可以促进西进运动的草原植物整个第四纪来自亚洲18,21。
来自亚洲的地中海盆地的殖民化可能发生在不同的路线14,25,26。类群从寒冷的亚洲大草原将会扩大到地中海盆地的北部弧和中欧,殖民开放栖息地,例如潘诺尼亚的地区和亚高山带的环境11,27。另一方面,thermophilous类群(如从阿拉伯半岛)会迁移到非洲的北部边缘26,28。并行迁移和扩张或收缩的草原植物群的范围11、物种形成过程3,8,28,29日。几个类群在草原还non-steppe亚裔多元化环境的西进运动在地中海盆地在晚新生代15,28,30.。
最特色的元素之一Irano-Turanian和地中海植物蒺藜(菊科部落Cardueae;cf。31日,32。Cardueae最大的部落之一的家庭约2500种33,34大部分的亚族和一些非常speciose属等矢车菊属,Cirsium,Cousinia,Jurinea,和光学表现出多样性中心在亚洲35,36,37,38。最近约会的分析显示,它们起源于西亚与扩张和辐射整个新生代地中海和亚洲中部37。的发展史和分布族Cardueae已经彻底研究,但是如此大的部落提供了多个例子值得深入研究,其中一个是属Rhaponticoides(亚族Centaureinae)。
根据最新的修订,Rhaponticoides由36个物种,狭窄的流行或广泛的分类单元从蒙古的大草原到伊比利亚半岛和北非33,39,40,41;无花果。1。然而,这个数字应该谨慎对待,因为它肯定是一个高估(见下文抽样Matherial和方法的细节)。从系统的角度来看,Rhaponticoides被认为是一个孤儿属Centaureinae因为特别孤立在亚族和我们忽略任何潜在的身份相对还是妹妹属35,37。尽管生物地理的兴趣42,我们缺乏一个time-calibrated生物地理学研究揭示过程潜在的电流分布属和揭示其祖先的区域以及迁徙路线在地中海盆地和物种形成事件。同样,一个全面的属的系统发育研究基于分子数据可能会支持或反驳的最新分类的建议41以及之前的形态学、孢粉学的和karyological研究39,40,43,44,45,46。

属的地理分布和抽样。每个人口的数量是指表1。这个数字是由使用http://maps-for-free.com/和Adobe Illustrator_26.5。
一些植物区系的生物地理学研究基于分子数据元素以惊人的析取在地中海盆地和附近的地区,如北极高山植物或兰德植物显示,观察到chorological模式是由更复杂的过程比由古典历史生物地理学提供7,47,48,49。观察到的分布和物种形成模式的草原在地中海盆地,研究比较,更不用说,通常通过调用Messinian模型来解决14,19。到目前为止,很少有现代生物地理学研究显式或隐式地Messinian模型进行测试。他们通常为场景提供事件和过程在不同的时间框架内根据类群进行了研究15,28,50。
在这种背景下,本研究旨在有助于更好地理解迁徙和多元化进程的当前chorological亚洲草原植物类群的相关模式。我们使用两个核区(ETS)和两个质体区域(rpl32-trnLUAG和ycf3-trnS)和贝叶斯推理进行约会,祖先的重建,和散布分析以下具体目标:(1)阐明的系统发育关系Rhaponticoides物种;(2)重建历史生物地理学使用约会和祖先的地区重建分析和测试如果属真正的地理起源与亚洲大草原;(3)重绘的扩张Rhaponticoides通过欧亚大草原和地中海盆地和重建途径和司机通过欧亚大陆和北非的扩张。
结果
的发展史Rhaponticoides
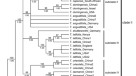
种Rhaponticoides放置在两核发展史moderately-supported演化支,配合双重属的分布(图2)。最speciose Irano-Turanian进化枝,包含所有的代表Irano-Turanian土耳其的一部分,亚美尼亚,和伊朗,加上物种从欧亚大陆的草原。地中海进化枝从地中海类群安纳托利亚的一部分,包括南巴尔干半岛,斜体和伊比利亚半岛。

50%的多数决定原则共识树通过贝叶斯组合核数据集的分析,表明支持演化支。数字发生以上分支机构后验概率(PP低于0.6不是如图所示)。引导可能性值图下面的分支(BS)没有显示低于60%。大写字母的名字后物种对应国家原产地物种与多个样本(见表1)。BG保加利亚,DZ阿尔及利亚,西文西班牙,FR法国,在印度,PT葡萄牙,TJ塔吉克斯坦、TR土耳其。
核(图。2(图)和质体DNA。3)phylogrames基本上是重合的,硬不一致:Rhaponticoides africana和Rh。fraylensis被放置在地中海进化枝的核数据(无花果。2)和Irano-Turanian进化枝的质体发展史(无花果。3)。同样的,Rh。iconiensis,Rh。gokceoglui,Rh。amasiensis和Rh。phytiae被放置在Irano-Turanian进化枝核发展史(无花果。2)而相同的四个物种是位于地中海进化枝在质体树中,尽管只有的位置Rh。gokceoglui支持(无花果。3)。在任何情况下,低分辨率的质体发展史的结果几乎不受支持的地中海进化枝。

50%的多数决定原则共识树通过贝叶斯质体组合数据集的分析,表明支持演化支。数字发生以上分支机构后验概率(PP低于0.6不是如图所示)。引导可能性值图下面的分支(BS)没有显示低于60%。大写字母的名字后物种对应国家原产地物种与多个样本(见表1)。BG保加利亚,DZ阿尔及利亚,西文西班牙,FR法国,在印度,PT葡萄牙,TJ塔吉克斯坦、TR土耳其。
生物地理学在time-calibrated框架
BayArea + J模型重建的祖先区域表现最好Rhaponticoides(表S2)。最后time-calibrated生物地理的模型显示,土耳其是最合理的祖先的面积Rhaponticoides中间的中新世马(12)。属分化两个地理线,中新世末Irano-Turanian和地中海演化支,马(8)。又两个分支多元化Miocene-Pliocene过渡(5 Ma)和显示一个辐射过程Pliocene-Quaternary过渡和整个第四纪(无花果。4、表3)。

约会和分子生物地理的分析。最大进化枝信誉树从放松分子钟分析生死与指数分布和物种形成过程和ETS序列的野兽。数字参考节点(表支持3)。只对节点支持饼图(如图所示,表中3)的相对概率反映每个区域或地区被祖先的组合,根据祖传的地区重建基于BioGeoBEARS BayArea + J模型实现。字母对应的祖先区域或结合区域上方的数字表示。这个数字是由使用http://maps-for-free.com/、QGIS_3.4 Inkscape_0.92.3。
多元化进程带来不同的扩张过程对西方和中亚,一方面,对地中海盆地。这种变化发生在不同的血统和在不同的日期。Irano-Turanian草原行起源于土耳其的草原环境,即。安纳托利亚高原(图中红色区域。4)和多样化(如。Rh。amasiensis)(节点在图6和10。4)。这条线也扩大对伊朗和高加索(节点7和8,路线在图1。4、表3),引发了伊朗物种类群复杂(Rh。lachnopus)和高加索集团(Rh。hajastana)。也源自相同的分支Rh。阿尔/ ruthenica复杂,集出现的人口从西印度到阿尔卑斯山伊比利亚半岛和地中海盆地。这些欧洲人(黄色和绿色的区域在无花果。4)代表向西迁移跟踪(路线在图1 b。4在地中海盆地北部边缘的)。
地中海线(节点在图11。4)也表现了亚洲不可告人的多样化的物种起源。这条线和大草原在土耳其一个祖先区域(图中红色区域。4),是第一个进化枝(节点在图11和12。4在安纳托利亚与地中海气候(南部)Rh。hierroi和Rh。mykalea)。这个地中海线达到巴尔干半岛(Rh。amplifolia和Rh。wagenitziana)和南意大利(Rh。calabrica和Rh。centaurium)构成Balkan-Greek-Italic血统(节点13、14和16、路线在图2。4)。地中海线也扩展到整个地中海的栖息地从南意大利(Rh。非洲象)到北非(Rh。非洲象,Rh。eriosiphon[presumibly灭绝,苏珊娜per。通讯。])伊比利亚半岛(Rh。非洲象和Rh。fraylensis)(节点17日路线在图2。4)。
讨论
的起源Rhaponticoides、速度和位置。草原植物和植被出现在地中海盆地早第三纪以来(包括西欧)并获得整个新第三纪的相关性,特别是在中新世10,20.。的地理起源的主要草原血统,殖民统治地中海盆地是亚洲中部和西部15,30.,51这是同样的情况Rhaponticoides(无花果。4),因为土耳其是最可能的祖先区域。土耳其的安纳托利亚高原景观为主,属于Irano-Turanian地区15,52和地中海带位于安纳托利亚半岛的南部和西部。
安纳托利亚高原,位于欧洲的十字路口,亚洲和非洲,受益的多样性三个热点地区:东地中海,Iran-Anatolia和高加索地区29日,50。也是一个会议地点和疏散走廊为不同的血统,起源于亚洲和欧洲和北非殖民地在新生代15,53,其中亚族Centaureinae哪Rhaponticoides属于37。此外,安纳托利亚高原展示高水平的特有现象和丰富的生物多样性可能由一个复杂的古地理历史戏剧性的新生代期间的地形和气候变化54,55。
的起源Rhaponticoides应放置在中中新世的建议不同类群的维管植物Irano-Turanian起源50,56从菊科植物和许多团体33,34。最多样化在Cardueae事件,特别是在亚族Centaureinae复发相关之间的连接和隔离集安纳托利亚高原和地中海盆地中新世37。反过来,这些情节都与环境变化在Irano-Turanian和地中海等地区气候变化,通常倾向于冷却和aridification,和碰撞的构造板块和随后的坑底隆起的高原和山脉组成接近安纳托利亚的人,例如,扎格罗斯29日,50。
多样化的Rhaponticoides。的多元化Rhaponticoides发生在晚新生代(无花果。4为其他维管植物血统)作为描述57。它开始在中新世末后续Miocene-Pliocene辐射事件和Pliocene-Quaternary转换(无花果。4)。第一个发生在中新世末和多元化Miocene-Pliocene过渡尤其是隐含色散。注册在其他草原Irano-Turanian类群25,49,Rhaponticoides目前草原血统扩大向东和起源物种生长在伊朗Rh。lachnopus。同时,属地中海线表明,起源于土耳其的地中海地区,而迁移到西部到达欧洲和西北非洲西南部。这些分散和多元化的事件与这些推断其他草原旱生植物出现在地中海盆地等远征28,Acantholimon29日,Haplophyllum15。扩大Irano-Turanian旱生植物已被极端aridification青睐和部分干燥的地中海Messinian危机期间,这将导致大陆连接,即。,走廊向西方地中海盆地的结束。因此,早期的多样化Rhaponticoides适合Messinian模型14。
Messinian模型兼容反过来后分散和多元化进程发生在Pliocene-Quaternary过渡和第四纪,发病可能与地中海气候8,58也与气候和海平面振荡与冰期相关联21,28,29日。的主要血统Rhaponticoides这以前起源于Miocene-Pliocene过渡经验丰富的多样化和地中海盆地和西方殖民亚洲。地中海线进化可能来自大草原的物种发生在地中海带安纳托利亚(如。Rh。amasiensis,Rh。hierroi,Rh。mykalea)和亚美尼亚(Rh。hajastana),然后迁移到地中海东部和中部(巴尔干半岛和意大利,例如,Rh。amplifolia, Rh。calabrica,Rh。centaurium,Rh。wagenitziana)。此外,在大草原上,一些物种出现,取得了广泛和分离的分布通过多种原始的碎片或长距离传播事件59。是这样的Rh。阿尔/ ruthenica复杂与间断分布在不同的山脉:从在中亚天山到高加索,巴尔干半岛,欧洲阿尔卑斯山脉和伊比利亚半岛41,42。取得了类似的结果在其他草原类群起源于Irano-Turanian地区28,29日,51。
考虑到环境的变化在新生代后期,地理分隔可以解释,至少部分,血统和物种的起源Rhaponticoides。然而,物种形成传播事件似乎更多的频率比地理分隔在草原分类单元位于地中海和Irano-Turanian区域15,29日,30.,51。在我们的分析中,BayArea + J被选为最佳的模型,这个模型包括分枝进化和共存性以及再生和分散而地理分隔没有相关性60。Rhaponticoides物种的草原和地中海线起源于生物地理学特征的地区和安纳托利亚高原等复杂环境历史,哪个港口多个特有种54,55。范围扩张被走廊青睐通过Messinian危机等事件15,29日。
质体和核数据之间的不一致在伊比利亚发现Rh。fraylensis和Rh。非洲象(无花果。2和3)是非常直观的起源Irano-Turanian池内的地中海线:两个物种出现在质体发展史中嵌套Irano-Turanian进化枝。通常的解释不一致是古代杂交和随后的质体捕获61年,62年。在我们的例子中,最合理的假设表明的祖先Rh。非洲象和Rh。fraylensis收购了Irano-Turanian叶绿体基因渗入的演化支在安纳托利亚之间的接触区。质体和核的发展史之间的其他不一致(例如,Rh。amasiensis,Rh gokceoglui,Rh。iconiensis和Rh。phytiae)可以减少到只有一个支持不一致Rh。gokceoglui自的位置Rh。amasiensis,Rh。iconiensis和Rh。phytiae不支持在质体树(PP = 0.76, b = 63%,无花果。3)。冲突的位置Rh。gokceoglui还应该解释为一个古老的杂交和质体捕获,这是一个常见的fenomenon部落Cardueae吗33,36。
对地中海盆地迁徙路线。到目前为止,两个路线通常报告Irano-Turanian草原植物目前位于Mediterranenan盆地。一些类群遵循了“北方路线”包括山脉和/或草原的南欧和地中海北部边缘的11,27。相比之下,其他类群跟踪通过北非“南部路线”,地中海岛屿,最终通过陆地出现在新第三纪和第四纪的干旱危机26,28。Rhaponticoides由血统,比赛路线和反映了生物地理的模式(路线在图1 b和2。4)。
关于北路线(路线在图1 b。4),大草原,起源Rh。阿尔/ ruthenica复杂到地中海盆地的西部边缘(伊比利亚半岛)通过巴尔干半岛和阿尔卑斯山脉,离开在渣土relictic的数量Rh。阿尔作为里程碑42。这条线还包括从西部和中亚大草原物种分布集中在Irano-Turanian地区到达伊朗南部的草原(在我们分析的代表Rh。lachnopus)(图。4)。至于南方路线(路线在图2。4),地中海线集成了一系列窄分布的物种,其中许多来自地中海安纳托利亚的一部分41,46向西延伸到北非和伊比利亚半岛(无花果。4)。
共存的途径和两个lines-steppe和地中海的到来伊比利亚半岛匹配的情况矢车菊属教派。Acrocentron(Cass)。直流。25。的途径矢车菊属从西西里到西班牙是被一些站作为垫脚石的残遗种,即(从东到西)c . tauromenitana,c . carolipauana,c . clementei。相比之下,在南方的踏脚石的途径Rhaponticoides由只有残遗,孤立的单个物种的数量,即Rhaponticoides africana,用小站在西西里,北非,南部和西北伊比利亚。非凡的旅程Rh。非洲象在加利西亚(西班牙西北部)结束,一些残余的人口几乎靠沿海小石子63年。
分类的影响。这篇文章的目的不是分类,但是我们的结果表明,该属的分类学远非众所周知的。的infrageneric分类法Rhaponticoides研究了很久但主要基于形态学标准。最详尽的和最近的工作41提出了一种分段分类属的几乎所有物种是一个部分或分段,同意39。然而,我们的研究结果揭示差异单系统的一些部分和物种。部分Ruthenicae和Aralocaspicae不是单元,它们包括物种恢复到不同的演化支无关。这样的不一致性可能出现的物种界定39。例如,分离的形态学证据Rh。ruthenica从Rh。阿尔非常弱,只是基于一些营养字符41而我们的系统发育结果证实,这种隔离缺乏支持从分子证据(无花果。2)。
在同一条线上,许多分类实体在物种水平相当可疑的价值在我们看来是描述在宽范围内Rh。ruthenica。支持这一观点,我们的结果Rh。razdorskyi这些隔离物种之一,分组Rh。ruthenica。在相同的部分Ruthenicae的独立Rh。linaresii在我们分析的代表,瓦伦西亚的人口Rh。阿尔和持续的41已经拒绝了所有的修订42,64年合并成Rh。阿尔证实了我们的结果。同样的,Rh。carrisoi被列为单独的物种41但它实际上是一个同义词Rh。fraylensis64年。最后,Rh。centaurium和Rh。calabrica被认为是同义词65年根据我们的结果,是非常可能的。相反,物种会非常狭隘的连接Rh。wagenitziana从巴尔干半岛45。
总之,全球财富和一些界定infrageneric实体,部分和物种Rhaponticoides需要一个深,严重的修订,应该综合分类的框架内进行66年。我们建议将更广泛的代表类群和phylogenomic的使用方法已经用于Cardueae37。
结束语
属设置其起源的历史生物地理学的Irano-Turanian部分土耳其安纳托利亚高原中中新世。属经验不同的多元化发展和向西和向东扩张不仅Messinian盐度相关的危机还在Pliocene-Quaternary不可告人的环境变化和第四纪时期。Rhaponticoides像其他Irano-turanian草原类群,殖民统治地中海盆地在不同的日期。然而,在最前面研究的分类单元相比,属Rhaponticoides迁移和多样化的地中海盆地后两种不同的路线。在建设的系统发育关系Rhaponticoides揭示了一个全面综合的迫切必要性研究属解决一些infrageneric类群的界定。
材料和方法
方法符合当地和国家的指导方针。
抽样
抽样旨在覆盖所有的区域和细胞核多样化:属高加索,伊朗,巴尔干半岛,伊比利亚半岛、意大利和西西里,北非和欧亚大陆的草原,在印度地区的极端(无花果。1)。重点提出了在土耳其,根据峰值显示属物种44。大多数物种上市39,41缺少我们抽样怀疑将是广泛的Rh。ruthenica;事实上,属的物种的总数可能是接近25 - 30,这使得我们的报道60 - 72%的属。针对不同治疗一些类群和一些其他的非常大的区域,包括我们几个的数量Rhaponticoides africana,Rh。阿尔和Rh。ruthenica。抽样总数26 18物种的种群。系统发育分析,外围集团被选中37包括两个属的物种Klasea卡斯。,namelyKlasea coriacea(费。& c.a最大经济产量。直流交货)。Holub和k . serratuloides(直流)。Greuter & Wagenitz(数据集1和2)。测定分析,添加了其他外围集团(见下文,数据集3)。材料和凭证的起源在表表示1。
DNA提取和放大
总基因组DNA提取硅gel-dried一植物人口的组织。提取的DNA的CTAB方法67年的修改68年包括三个清洗步骤与山梨糖醇缓冲区。ETS,其rpl32-trnLUAG,和ycf3-trnS地区被聚合酶链反应(PCR)放大。核区域的扩增引物是ITS1和ITS469年它和ETS1F70年和18集71年ETS的地区。质体的rpl32-trnLUAG地区,我们使用rpl32-F底漆和trnLUAG随着反向引物72年。质体的ycf3-trnS地区,我们使用SP43122F作为正向引物和SP44097R反向引物73年。PCR反应条件下进行了详细74年。放大DNA片段测序在ABI 3730 xl分析器(美国应用生物系统公司,培育城市,CA)在Macrogen Inc .制造商的协议后,韩国。
系统发育分析
核苷酸序列编辑使用Bioedit v7.0.5.375年和对齐视觉序列两两比较76年。在其基本数据,ETS,rpl32-trnLUAG,和ycf3-trnS矩阵表是可用的S3。其加ETS矩阵长1594个基点(数据集1)和对齐的质体矩阵长1821个基点(数据集2)。可能性分析的数据集是由利用PAUP v4.0启发式搜索77年用为分支交换角色指定为无序和无关紧要的。可能性标准将HKY85 PAUP(默认选项)。引导分析78年进行了使用100复制启发式搜索的默认选项。节间的引导(BS)值> 75%被认为是具有统计学意义。
贝叶斯推理的两个数据集计算使用MrBayes v3.2.679年。最好的可用模型的贝叶斯估计所需的分子进化发展史选择使用Akaike信息标准(AIC)对数据集MrModeltest v2.2中实现软件80年。最好的拟合模型GTR + G +我对数据集。贝叶斯推理分析了随机树开始,运行40×106代两个独立运行的四个Metropolis-coupled链。我们救了一个每1000代,导致40000采样树。数据从1000年第一个采样树被丢弃的“老化”时期,在确认对数似值稳定在1000采样树。运行的平稳性和之间的融合与示踪v.1.5.0检查81年。节间和后验概率(PP) > 0.95被认为是具有统计学意义。
测定分析
分歧时间估计使用nrDNA (ETS)及其序列在一个矩阵组织一次到达每个派系(数据集3,表的分类单元1)。外围集团包括Klasea coriacea,k . serratuloides(如数据集1)优先Plagiobasis centauroidesSchrenk,Leuzea acaulis(l)Holub。,l . conifera(l)直流。通过使用野兽v.1.8.4约会分析。四个单元组中定义的美丽女孩v.1.8.4(包括在野兽包):(i)所有物种的数据集2 (2)Plagiobasis centauroides(3)Klasea+Leuzea进化枝,(iv)和Rhaponticoides属。这四组同时也作为二次校准点实现基于先前phylogenomic研究集中在Cardueae部落37,见表2。
获得最佳time-calibrated phylogram、模型与对数正态分布放松严格的和不相关的时钟是两个不同的物种形成树模型下运行,圣诞和生灭82年,83年。对数正态分布和指数的选择也测试模型与对数正态分布的情况下轻松的时钟(见表S1)。六个结果野兽模型运行四个独立链5000万代,抽样每1000代。他们的收敛性是评估确认所有参数达到了平稳性和充分有效的样本大小(> 200)在所有聚合使用示踪v1.7运行81年。模型和复制都是运行在颁发科学网关84年。
六6根据各自不同的野兽模型比较日志的价值观(企业)边际似然估计得到path-sampling (PS)和方法(SS)85年实现野兽v.1.8.4 S1(见表)。PS和SS日志值估计有100路径步骤,106代的链长和可能保存每1000代。毫升的生成的日志值平均四个复制运行生成一个PS和SS价值为每个模型。日志的平均获得值毫升的所有假设都是排名,和贝叶斯因子(BF)然后使用修改引入的计算86年(即。,twice the difference between the harmonic mean likelihoods of the two models). Values for 2BF those are greater than 2, 6, and 10 indicate positive, strong, and decisive support, respectively, for the generic hypothesis with minor marginal likelihood.
最好的模型与决定性的支持是一个轻松的时钟生死与指数分布和物种形成过程(表S1)。丢弃老化步骤之后(25%)、树文件从所选的四个独立的运行模型结合使用LogCombiner 1.8.4和由此产生的最大进化枝信誉(MCC)树中总结TreeAnnotator 1.8.4 (https://beast.community/2016-06-17_BEAST_v1.8.4_released.html)和无花果树从v.1.4.2 (https://github.com/rambaut/figtree/releases/tag/v1.4.4)。
祖先的面积估计
我们推断属的起源Rhaponticoides及其可能的途径扩张和物种形成的种间系统发育关系支持time-calibrated树受到生物地理学分析,BioGeoBEARS87年。我们定义9地理区域为基础,主要物种的丰富性和地方的特性Rhaponticoides属(无花果。4,如上图所示)。time-calibrated树产生的贝叶斯推理的野兽被用作输入文件估计祖先的概率范围(数据集3)。BioGeoBEARS计算祖先州内部节点的最大似然估计模型之间的转换地理范围以及系统发育分支作为时间的函数。BioGeoBEARS包括六个不同的生物地理学模型(12月,12月+ J,女主角,女主角+ J, BAYAREA-LIKE, BAYAREA-LIKE + J) BioGeoBEARS R包中实现87年。所有的模型需要分散(d)和灭绝(e)作为自由参数。三个模型还包含一个额外的参数“j”(+ j)拥抱founder-event物种形成87年但是12月+ J丢弃根据最近的批评88年。BioGeoBEARS收益率祖先州内部节点的最大似然估计模型之间的转换地理范围以及系统发育分支作为时间的函数。六个生物地理学模型比较适合的使用价值和可能性Akaike信息准则(AIC)。因此,在我们的研究中,BayArea + J模型(表中表现最好S2)。
伦理语句
所有样本包含在各自的研究来自标本采样允许从国家政府和植物标本室(ATHU、国家和大学Kapodistrian希腊雅典;KNYA科尼亚标本、土耳其;西班牙巴塞罗那BC,植物研究所;CIEF Servicio·德·维达西尔维斯特,Generalitat Valenciana,西班牙;E,苏格兰爱丁堡皇家植物园;HLUC,意大利degli某黛拉巴斯利卡塔、意大利;惠,艺术学校botanique国家mediterraneen de Porquerolles、法国;GAZI大学GAZI土耳其;塞夫、塞维利亚大学、西班牙;SOM,生物多样性和生态系统研究所的植物标本,保加利亚科学院Bulgary)。
数据可用性
数据集1、2和3可以在线访问https://www.ibb.csic.es/public/。凭证标本数据和基因库加入数字序列表中描述1。
引用
乌鸦,p h .灾难性的选择和土壤特有现象。进化18,336 - 338。https://doi.org/10.2307/2406409(1964)。
他们批判,J。,Aronson, J. & Boeuf, G.地中海地区(牛津大学出版社,2010年)。
Rundel, p W。et al。火和植物多样化在地中海气候地区。科学前沿Pl。9,851年。https://doi.org/10.3389/fpls.2018.00851(2018)。
乌鸦,p . h .地中海组成的进化。在地中海类型生态系统:起源和结构(eds di Castri f . &穆尼,h·a .) 213 - 224 (Springer, 1973)。https://doi.org/10.1007/978 - 3 - 642 - 65520 - 3 - _14。
巴伦,E。et al。伊比利亚半岛的新生代植物:合成。启Palaeobot。Palynol。162年,382 - 402。https://doi.org/10.1016/j.revpalbo.2009.11.007(2010)。
阿克塞尔罗德,d .进化和生物地理学Madrean-Tethyan硬叶植物。安。密苏里州的机器人。加尔省。62年,280 - 334 (1975)。
尼托猫,g模式和过程在植物phylogeography在地中海盆地:复习一下。教谕。Pl,生态。另一个星球。系统。16,265 - 278。https://doi.org/10.1016/j.ppees.2014.07.002(2014)。
巴尔加斯,P。,Fernández-Mazuecos, M. & Heleno, R. Phylogenetic evidence for a Miocene origin of Mediterranean lineages: Species diversity, reproductive traits and geographical isolation.Pl,杂志。20.,157 - 165。https://doi.org/10.1111/plb.12626(2018)。
Le Houerou h . n .撒哈拉以北干旱steppeland生物地理学。j .干旱的环境。48,103 - 128。https://doi.org/10.1006/jare.2000.0679(2001)。
Hurka, H。et al。欧亚草原地带:现状、起源和演化历史。Turczaninowia225 - 71。https://doi.org/10.14258/turczaninowia.22.3.1(2019)。
Kajtoch,Łet al。Phylogeographic模式在欧洲中部东部草原物种:回顾和对保护的影响。Biodivers。Conserv。25,2309 - 2339。https://doi.org/10.1007/s10531 - 016 - 1065 - 2(2016)。
SainzOllero, h。全世界的大草原:概述和伊比利亚半岛上的重点。在草原生态系统:生物多样性、管理和恢复(eds莫拉莱斯,m & Traba j .) 1-26(新星出版社,2013)。
Wesche, K。et al。古北区的草原生态区:一个新的合成。Biodivers。Conserv。25,2197 - 2231。https://doi.org/10.1007/s10531 - 016 - 1214 - 7(2016)。
Bocquet G。,Widler, B. & Kiefer, H. The Messinian model: A new outlook for the floristics and systematics of the Mediterranean area.Candollea33,269 - 287 (1978)。
Manafzadeh, S。,Salvo, G. & Conti, E. A tale of migrations from east to west: The Irano-Turanian floristic region as a source of Mediterranean xerophytes.j . Biogeogr。41,366 - 379。https://doi.org/10.1111/jbi.12185(2014)。
布拉姆韦尔,加那利群岛的流行菌群:分布、关系和植物地理学。在生物地理学和生态学在加那利群岛(ed, Kunkel) 207 - 239 (w . Yunk出版社,1976)。
Vai g . b .超过半个世纪的Messinian盐度危机。波尔。青烟。分钟。127年,615 - 632 (2017)。
Yaprak, a & Kadereit g . Microcnemum珊瑚状的(Chenopodiaceae-Salicornioideae):种内东西析取的一个例子在地中海地区。肛门Jard。机器人。马德里65年,415 - 426。https://doi.org/10.3989/ajbm.2008.v65.i2.303(2008)。
Ribera, i & Blasco-Zumeta j .生物地理学Monegros地区草原昆虫之间的联系(西班牙阿拉贡,NE),东地中海和中亚。j . Biogeogr。25,969 - 986。https://doi.org/10.1046/j.1365-2699.1998.00226.x(1998)。
Jimenez-Moreno, g . &往下,j.p.中产中新世纬向气候梯度在西欧:证据从花粉记录。Palaeogeogr。Palaeoclimatol。Palaeoecol。253年,208 - 225。https://doi.org/10.1016/j.palaeo.2007.03.040(2007)。
Seidl,。et al。系统学和生物地理学的更新世全北区的草原和半荒漠植物藜属植物Krascheninnikovia驼绒藜。植物区系262年,151504年。https://doi.org/10.1016/j.flora.2019.151504(2019)。
吴,H。,Guiot, J., Brewer, S. & Guo, Z. Climatic changes in Eurasia and Africa at the last glacial maximum and mid-Holocene: Reconstruction from pollen data using inverse vegetation modelling.爬。直流发电机。29日,211 - 229 (2007)。
休伊特,g .冰河时代:物种的分布和演化。在地球上的进化(埃德•罗斯柴尔德。l . j . &李斯特,a . m .) 339 - 361(学术出版社,2003年)。https://doi.org/10.1016/b978 - 012598655 - 7/50045 - 8所示。
兰贝克,K。Rouby, H。,Purcell, A., Sun, Y. & Sambridge, M. Sea level and global ice volumes from the Last Glacial Maximum to the Holocene.Proc。国家的。学会科学。美国111年,15296 - 15303。https://doi.org/10.1073/pnas.1411762111(2014)。
字体,M。,Garcia-Jacas, N。,Vilatersana, R., Roquet, C. & Susanna, A. Evolution and biogeography of矢车菊属部分Acrocentron推断出从核质体DNA序列分析。安。机器人。103年,985 - 997。https://doi.org/10.1093/aob/mcp022(2009)。
Perez-Collazos E。,Sanchez-Gómez, P., Jiménez, J. F. & Catalán, P. The phylogeographical history of the Iberian steppe plant Ferula loscosii (Apiaceae): A test of the abundant-centre hypothesis.摩尔。生态。18,848 - 861。https://doi.org/10.1111/j.1365 - 294 x.2008.04060.x(2009)。
Hensen,我。et al。低的遗传变异和强大的分化孤立人群罕见的大草原上的草大针capillatal在欧洲中部。Pl,杂志。12,526 - 536。https://doi.org/10.1111/j.1438-8677.2009.00227.x(2010)。
Lauterbach, M。,Veranso-Libalah, M. C., Sukhorukov, A. & Kadereit, G. Biogeography of the xerophytic genus远征l .(藜科)。生态。另一个星球。9,3539 - 3552。https://doi.org/10.1002/ece3.4987(2019)。
Moharrek F。,Sanmartin,我。,Kazempour-Osaloo, S. & Nieto Feliner, G. Morphological innovations and vast extensions of mountain habitats triggered rapid diversification within the species-rich Irano-Turanian genusAcantholimon(Plumbaginaceae)。前面。麝猫。9,698年。https://doi.org/10.3389/fgene.2018.00698(2019)。
将球击出,C。et al。与贝叶斯方法重建的历史桔梗科分子约会和Dispersal-Vicariance分析。摩尔。Phylogen。另一个星球。52,575 - 587。https://doi.org/10.1016/j.ympev.2009.05.014(2009)。
Quezel, p .地中海和非洲撒哈拉的菌群分析。安。密苏里州的机器人。加尔省。65年,479 - 534。https://doi.org/10.2307/2398860(1978)。
Takhtajan,。植物区系地区的世界(加州大学出版社,1986年)。
苏珊娜,a & Garcia-Jacas:部落Cardueae。在开花植物(eds Kadereit j . w . &杰弗里·c·g .) 123 - 146 (Springer, 2007)。
苏珊娜,a & Garcia-Jacas: Cardueae (Carduoideae)。在菊科植物的分类学,进化和生物地理学(eds恐慌,诉。et al。)293 - 213 (IAPT, 2009)。
Garcia-Jacas, N。苏珊娜,一个。,Garnatje, T. & Vilatersana, R. Generic delimitation and phylogeny of the subtribe Centaurineae (Asteraceae): A combined nuclear and choloroplast DNA Analysis.安。机器人。87年,503 - 515 (2001)。
Lopez-Vinyallonga, S。et al。系统学和进化的Arctium-CousiniaCardueae-Carduinae复杂(菊科)。分类单元58,153 - 171。https://doi.org/10.1002/tax.581016(2009)。
Herrando-Moraira, S。et al。核和质体DNA发展史的部落Cardueae(菊科)Hyb-Seq数据:一个新的subtribal分类多样化和时间框架。摩尔。Phylogenet。另一个星球。137年,313 - 332。https://doi.org/10.1016/j.ympev.2019.05.001(2019)。
Ackerfield, J。et al。一个棘手的难题:通用的界定Carduus-Cirsium组(菊科:Cardueae: Carduinae)。分类单元69年,715 - 738。https://doi.org/10.1002/tax.12288(2020)。
Agababian, M。矢车菊属subg。矢车菊属(菊科):界定和分配的部分或细节。Lagascalia19,889 - 902 (1997)。
Puntillo, d &佩鲁齐l .新物种RhaponticoidesVaill。从意大利南部(菊科)。叶形线Geobot。44,191 - 197。https://doi.org/10.1007/s12224 - 009 - 9037 - x(2009)。
Negaresh, k属的分类Rhaponticoides(菊科Cardueae)。Phytotaxa422年,9-32。https://doi.org/10.11646/phytotaxa.422.1.2(2019)。
苏珊娜,a . Mapa 64:矢车菊属阿尔l在Asientos对位联合国Atlas Corologico de la植物西方(ed, FernandezCasas j .) 17 - 20 (Fontqueria, 1988)。
Agababian m & Goukasian a诉的细胞核学亚美尼亚的代表矢车菊属亚属矢车菊属(菊科)。Thaiszia4,171 - 173 (1994)。
Ghukasyan, a . & Hayrapetyan a . m .孢粉学的和caryological调查属的特有种RhaponticoidesVaill。(fam)。菊科)包含在亚美尼亚的红色植物的数据手册。Nat。学会科学。亚美尼亚代表。115年,156 - 162 (2015)。
棕褐色,K。,Bancheva, S., Vural, M. & Strid, A.矢车菊属wagenitziana(菊科:Centaureinae),从巴尔干半岛东部的一个新物种。叶绿醇。Balcan。1551-58 (2009)。
Uysal, T。et al。Karyomorphological研究五土耳其流行RhaponticoidesVaill。(菊科Cardueae)物种。Caryologia69年,207 - 214。https://doi.org/10.1080/00087114.2016.1169088(2016)。
Holderegger, r &雅培,r . j . Phylogeography极地高山虎耳草属oppositifolia基于cpDNA (Saxifragaceae)和一些相关类群及其序列变异。点。j .机器人。90年,931 - 936。https://doi.org/10.3732/ajb.90.6.931(2003)。
Sanmartin,我。,Anderson, C. L., Alarcón, M., Ronquist, F. & Aldasoro, J. J. Bayesian island biogeography in a continental setting: The Rand Flora case.医学杂志。列托人。6,703 - 707。https://doi.org/10.1098/rsbl.2010.0095(2010)。
Pokorny, L。et al。生活在边缘:兰德植物的时机和持续的在非洲aridification析取相等的。前面。麝猫。6,154年。https://doi.org/10.3389/fgene.2015.00154(2015)。
Mahmoudi, M。,Moharrek F。,Assadi, M. & Nieto Feliner, G. Biogeographic history and diversification patterns in the Irano-Turanian genusAcanthophyllumsl。(石竹科)。Pl Biosyst。155年,425 - 435。https://doi.org/10.1080/11263504.2020.1756974(2021)。
彼得森,。Harpke D。,Peterson, J., Harpke, A. & Peruzzi, L. A pre-Miocene Irano-Turanian cradle: Origin and diversification of the species-rich monocot genusGagea(百合科)。生态。另一个星球。9,5870 - 5890。https://doi.org/10.1002/ece3.5170(2019)。
Manafzadeh, S。,Staedler, Y. & Conti, E. Visions of the past and dreams of the future in the Orient: The Irano-Turanian region from classical botany to evolutionary studies.医学杂志。启剑桥市费罗斯。Soc。92年,1365 - 1388。https://doi.org/10.1111/brv.12287(2016)。
Jabbour F。Consolida和Aconitella是一年一度的进化枝的飞燕草(毛茛科),在地中海盆地和Irano-Turanian地区多元化。分类单元60,1029 - 1040。https://doi.org/10.1002/tax.604007(2011)。
戴维斯p h .分布模式在安纳托利亚与特定的特有现象。在亚洲西南部的植物(eds戴维斯,p . H。et al。)15-28(植物社会爱丁堡,1971)。
Noroozi, J。et al。在土耳其的特有现象,会议的三个全球生物多样性热点地区,基于三维管植物的各种各样的家庭。前面。生态。另一个星球。7,159年。https://doi.org/10.3389/fevo.2019.00159(2019)。
吴,告诫。et al。进化的亚洲内陆干旱带生物群:证据来自亚洲的多元化霸王属(蒺藜科)。《公共科学图书馆•综合》10,e0138697。https://doi.org/10.1371/journal.pone.0138697(2015)。
太阳,M。et al。最近的加速多元化Rosids发生在热带地区。Commun Nat。11,3333年。https://doi.org/10.1038/s41467 - 020 - 17116 - 5(2020)。
往下,j。起源和演化在欧洲地中海的植被和气候。自然307年,429 - 432 (1984)。
Ansell, S。et al。安纳托利亚山的重要性成为全球多样性的摇篮而阿尔,一个关键的极地高山物种。安。机器人。108年,241 - 252。https://doi.org/10.1093/aob/mcr134(2011)。
Matzke: j .概率历史生物地理学:新的founder-event物种形成模型,完善检测&化石允许提高精度和模型试验。前面。Biogeogr。5,242 - 248 (2013)。
万德尔,j . f . & Doyle, j。j系统不一致:窗口基因组和分子进化历史。在植物分子系统学II: DNA测序(eds Soltis d E。et al。)256 - 296 (Kluwer学术出版社,1998)。
林德,c . r . &里瑟博格,l . h .重建网状进化的模式植物。点。j .机器人。91年,1700 - 1708。https://doi.org/10.3732/ajb.91.10.1700(2004)。
苏珊娜,a . Mapa 510。矢车菊属非洲"林。在Asientos对位联合国atlas corologico de la植物西方(eds费尔南德斯卡萨斯,j . &加马拉r .) 206 - 208 (Fontqueria, 1993)。
Pujadas, A . j . & Devesa j . A。RhaponticoidesVaill。在植物Iberica(eds Devesa j。et al。(CSIC) 263 - 272, 2014)。
Bartolucci F。et al。Notulae意大利本土血管弗洛拉:3。意大利的植物学家3,29-48。https://doi.org/10.3897/italianbotanist.3.13200(2017)。
Miguez, M。,Martín-Bravo, S. & Jiménez-Mejías, P. Reconciling morphology and phylogeny allows an integrative taxonomic revision of the giant sedges of苔属植物部分Rhynchocystis(莎草科)。机器人。j·林恩。Soc。188年,34-58。https://doi.org/10.1093/botlinnean/boy040(2018)。
柯南道尔,j。j &迪克森·e·e·保护植物样品DNA限制性内切核酸酶分析。分类单元36,715 - 722 (1987)。
Tel-Zur, N。、Abbo年代。,Mysladbodksi, D. & Mizrahi, Y. Modified CTAB procedure for DNA isolation from epiphytic cacti of generaHylocereus和Selenicereus(仙人掌科)。Pl。摩尔。杂志。代表。17,249 - 254 (1919)。
白色,t·J。布鲁斯,T。,Lee, S. & Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. InPCR协议:指导方法和应用(eds英尼斯,m。et al。)315 - 322(学术出版社,1990年)。
林德,c R。,Goertzen, L. R., Vanden Heuvel, B., Francisco-Ortega, J. & Jansen, R. K. The complete external transcribed spacer of 18S–26S rDNA: Amplification and phylogenetic utility at low taxonomic levels in Asteraceae and closely allied families.摩尔。Phylogenet。另一个星球。14,285 - 303 (2000)。
鲍德温,b . g . &马科斯·s·系统实用程序的外部转录间隔区(ETS) 18 s-26s rDNA: ETS的同余及其树Calycadenia(菊科)。摩尔。Phylogenet。另一个星球。10,449 - 463 (1998)。
肖,J。,Lickey, E. B., Schilling, E. E. & Small, R. L. Comparison of whole chloroplast genome sequences to choose noncoding regions for phylogenetic studies in angiosperms: The Tortoise and The Hare III.点。j .机器人。94年,275 - 288 (2007)。
Hershkovitz, m·a .核糖体和叶绿体DNA证据表明西方美国马齿苋科在安第斯地区的多元化。Gayana机器人。63年13 - 74 (2006)。
Lopez-Alvarado, J。et al。岛屿作为进化谱系的十字路口:一个案例研究的矢车菊属教派矢车菊属(菊科)从撒丁岛(地中海盆地)。《公共科学图书馆•综合》15e0228776 (2020)。
大厅,t·A·BioEdit:一个用户友好的生物序列比对编辑和分析程序在Windows / NT 95/98。核酸协会。爵士。41,95 - 98 (1999)。
沃福德,d . l . &奥尔森g . j .发展史重建。在分子系统学(eds Hillis d m &莫里茨,c) 411 - 501(1990年Sinauer Associates)。
沃福德,d . L。使用吝啬PAUP * 4.0 b2:系统发育分析(Sinauer Associates 2002)。
Felsenstein, j .信心限制的发展史:使用引导的方法。进化39,783 - 791 (1985)。
Ronquist F。et al。MrBayes 3.2:贝叶斯系统推理效率和模型选择在模型空间。系统。医学杂志。61年,539 - 542 (2012)。
尼兰德,j·a·a·MrModel测试。2.2版。计算机程序作者分布(2004)。
兰姆伯特,一个。,Drummond, A. J., Xie, D., Baele, G. & Suchard, M. A. Posterior summarization in Bayesian phylogenetics using Tracer 1.7.系统。医学杂志。67年,901 - 904。https://doi.org/10.1093/sysbio/syy032(2018)。
圣诞,g .进化的数学理论,基于j·c·威利斯博士的结论,f·r·S。费罗斯。反式。B213年21 - 87。https://doi.org/10.1098/rstb.1925.0002(1925)。
Gernhard, t .条件重建过程。j理论的。医学杂志。253年,769 - 778。https://doi.org/10.1016/j.jtbi.2008.04.005(2008)。
米勒,m。,Pfeiffer, W. & Schwartz, T. Creating the cipres science gateway for inference of large phylogenetic trees. In:网关计算环境学报》研讨会(GCE),1 - 8 (2010)。
Baele G。,Li, W. L. S., Drummond, A. J., Suchard, M. A. & Lemey, P. Accurate model selection of relaxed molecular clocks in Bayesian phylogenetics.摩尔。杂志。另一个星球。30.,239 - 243。https://doi.org/10.1093/molbev/mss243(2013)。
Kass r . e . &阿布a .贝叶斯因子。j。中央集权。Assoc。90年,773 - 795 (1995)。
Matzke: j .模型选择在历史生物地理学显示,在岛演化支founder-event物种形成是一个至关重要的过程。系统。医学杂志。63年,951 - 970。https://doi.org/10.1093/sysbio/syu056(2014)。
Ree, r·h·& Sanmartin i概念和统计问题12月+ J founder-event物种形成模型及其与12月通过模型选择。j . Biogeogr。45,741 - 749。https://doi.org/10.1111/jbi.13173(2018)。
确认
由美国Bancheva贡献的材料(生物多样性和生态系统研究所、保加利亚科学院),t . Constantinidis(生态学和系统学、国家和Kapodistrian雅典大学),p . Ferrer-Gallego (CIEF-Wildlife服务,Generalitat Valenciana),诉高贵(艺术学校Botanique国家Mediterraneen de Porquerolles), l . Rosati(学校农业、林业和环境科学、意大利degli某黛拉巴斯利卡塔),m . Ekici (Gazi大学生物系,理学院安卡拉,土耳其),m . Sağıroğlu(艺术与科学学院生物学系,Sakarya大学Sakarya,土耳其)和BC和E植物标本室。我们也感谢基础图(项目编号110 t948)和S.U.BAP (Project Number 17401176), and the Catalan government (“Ajuts a grups consolidats” 2017-SGR1116) for financial support.
作者信息
作者和联系
贡献
M.B.,K.E., N.G.J., A.S., J.A.C. and T.U. designed the research. M.B., N.G.J., K.E., A.S. and T.U. sampled and selected the specimens for the molecular analyses. M.B. and N.G.J. processed the specimens and obtained the sequences. J.A.C., N.G.J. and A.S. performed the phylogenetical analyses and the dating and biogeographical analyses, and also prepared all tables and figures. J.A.C., N.G.J. and A.S. wrote a first draft of the manuscript and all the authors contributed to the revision of the final version of the manuscript.
相应的作者
道德声明
相互竞争的利益
作者宣称没有利益冲突。
额外的信息
出版商的注意
施普林格自然保持中立在发表关于司法主权地图和所属机构。
补充信息
权利和权限
开放获取本文是基于知识共享署名4.0国际许可,允许使用、共享、适应、分布和繁殖在任何媒介或格式,只要你给予适当的信贷原始作者(年代)和来源,提供一个链接到创作共用许可证,并指出如果变化。本文中的图片或其他第三方材料都包含在本文的创作共用许可证,除非另有说明在一个信用额度的材料。如果材料不包括在本文的创作共用许可证和用途是不允许按法定规定或超过允许的使用,您将需要获得直接从版权所有者的许可。查看本许可证的副本,访问http://creativecommons.org/licenses/by/4.0/。
关于这篇文章

引用这篇文章
Bozkurt, M。,Calleja Alarcón, J., Uysal, T.et al。生物地理学的Rhaponticoides在地中海植物,Irano-Turanian元素。Sci代表1222019 (2022)。https://doi.org/10.1038/s41598 - 022 - 24947 - 3
收到了:
接受:
发表:
DOI:https://doi.org/10.1038/s41598 - 022 - 24947 - 3
