
文摘
生殖分工是群居膜翅目昆虫的一个特点。雌性生殖皇后区或非生殖的工人。在蚂蚁,工人经常显示进一步任务专业化与变化相关联的规模和/或形态。因为女性polyphenism通常在环境控制下,它被认为是表观遗传机制(如DNA甲基化)发挥核心作用,因为他们调解gene-by-environment交互。刺激基因的甲基化表皮生长因子受体(表皮生长因子受体)确实是控制工人大小在一个高度多态蚁。然而,它仍然是未知的表皮生长因子受体甲基化也可以调节工人大小单型的物种。通过结合实验药理学和分子生物学,我们表明,工人与大小表皮生长因子受体甲基化在两个单型的蚂蚁。此外,我们功能表明,表皮生长因子受体信号影响工人的大小。这些结果表明,工人大小调节表皮生长因子受体甲基化在蚂蚁的机械化守恒的但仍未开发的单型的物种。
介绍
表型可塑性发生在一个单一的基因型编码多个不同的表型1。在所有生物体,它往往是高度自适应。通过改变他们的表型,以应对外部线索,生物对环境变化做出反应,可以提高生存和繁殖。一些最显著的表型可塑性的形式存在于社会膜翅目昆虫(蜜蜂,黄蜂,和蚂蚁),相同的基因型可以导致形态、生理、和行为不同的女性种姓:大,肥沃的皇后区,专门从事生殖和较小,通常无菌工人维护确保殖民地2。是否女性幼虫发育成生殖女王或非生殖工作者通常源于环境(例如,食品质量或数量)或社会(例如,女王和育存在/没有)线索,导致基线基因组表达自己不同的发展路线3,4,5。
不像蜜蜂和黄蜂,无数蚂蚁表现出高度的工人多态性,即殖民地的工人可以大小不同和/或形态6,7。工人大小变化存在沿着光谱,从射的,哪里有轻微的等距差异,二态性,有多种,不同的工人subcastes显示标志,非均衡的身体特征的差异。工人多态蚁科一再出现,表明尺度依赖的分工在工人健康促进殖民地8,9,10。
数十年的研究已经致力于理解工人的进化和维护多态性在蚂蚁。然而,它的遗传和发育起源只是开始被破译。在分子水平上,身体大小是由进化守恒的生长调节途径,如胰岛素/胰岛素样生长因子信号(IIS)11雷帕霉素靶(TOR)12和表皮生长因子受体(EGFR)信号13。饮食引发的信号通路,导致细胞发出信号级联,促进细胞生长、增殖和分化14。在昆虫中,结果是生长调节激素蜕皮激素的生产15和保幼激素(JH)11,13,16。然而,它仍然未知如何生成这些途径已成为调整工人大小相同的基因型变化。最近,有人提出,表观遗传机制,如DNA甲基化,在起作用,因为他们调解gene-by-environment交互,将环境信号转化为持久的基因表达的变化,无需修改DNA本身17,18,19。大量研究调查了DNA甲基化是否影响queen-worker种姓的决心。在蚂蚁20.,21和蜜蜂22,23,皇后区和工人有不同的DNA甲基化模式与微分相关基因表达和/或选择性剪接。在蜜蜂中,沉默的表达DNA-methyltransferase 3基因,负责新创DNA甲基化,使worker-destined幼虫更加queen-like24。然而,多个研究未能发现任何caste-specific甲基化在蚂蚁签名25、黄蜂26,蜜蜂27,28,29日。
据我们所知,一项研究探讨了DNA甲基化会如何影响工人大小变化社会膜翅目昆虫。在多态蚁Camponotus floridanus高度可变大小的工人,幼虫DNA甲基化可以调节成年工人大小。促生长基因表皮生长因子受体(表皮生长因子受体)是不同的甲基化,并表示在幼虫注定要成为大工人(即。(即,专业)与小工人。未成年人)29日。此外,成年工人大小可以通过药物改变幼虫DNA甲基化改变:当幼虫进行全基因组hypomethylation,他们成长为更大的工人。相比之下,hypermethylated幼虫发育成小工人。在这个物种中,甲基化状态的表皮生长因子受体生成的大多数工人阶级内部大小的差异30.。
在这里,我们表明,药理的改变表皮生长因子受体甲基化与工人大小两个单型的蚂蚁属于不同的亚科,阿根廷蚂蚁经由(亚科Dolichoderinae)和法老蚁Monomorium pharaonis(亚科切叶蚁亚科)(图。1),这表明工人大小调节表皮生长因子受体甲基化是机械化守恒的蚁科。

工人的大小分布经由和Monomorium pharaonis。工人的情节显示绝对频率不同的宽度。两个物种表现出单峰工人大小分布(即。,工人们是单型的)。图片:阿根廷蚂蚁:©菲尔·莱斯特,使用许可;m . pharaonis4.0:ЗeмлepoйкинCC冲锋队。
结果
我们第一次确认的工人阶级经由和Monomorium pharaonis单型的。工人大小分布是基于测量的成年工人从野外采样和实验室股票殖民地,分别(无花果。1)。
解决DNA甲基化的角色在工人在单型的蚂蚁大小的规定,我们喂养幼虫与药理hypomethylating代理5-Aza-2脱氧胞苷(5-Aza-dC)31日。在这两个阿根廷蚂蚁和m . pharaonis大小,成年工人也显著大于幼虫给定5-Aza-dC(图和控制解决方案。2;电子辅助表S2)。值得注意的是,5-Aza-dC也改变了形状大小分布的物种;分布出现倾斜大尺寸比控制阿根廷蚂蚁,但小工人大小m . pharaonis(无花果。2;电子辅助表S2)。
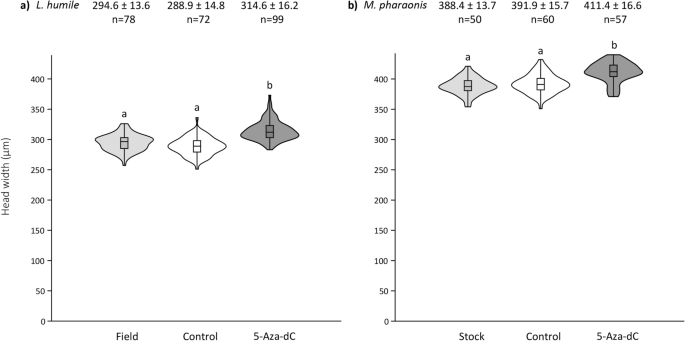
工人头上5-Aza-dC治疗宽度的影响。小提琴块(一个)经由和(b)Monomorium pharaoniscallow头宽度。工人被采样在球场上(阿根廷蚂蚁实验室殖民地()或形式m . pharaonis),或者从实验幼虫控制解决方案(1 M蔗糖)或者5-Aza-dC在第一次幼虫龄期。头宽度±SD和样本大小是上述每一个情节。框的中线表示中位数;框的上下边缘第一和第三个四分位数,分别。须反映极端值。盒子的形状是内核数据的密度分布。在小提琴上的字母表示差异在统计上有显著差异的工人头宽度(克鲁斯卡尔-沃利斯检验:p< 0.0001;邓恩的因果与Bonferroni调整测试:p< 0.0001)。
由于表皮生长因子受体信号控制经济增长中起着重要的作用32和表皮生长因子受体甲基化调节成年工人大小在一个多态蚁30.然后我们测试,如果治疗5-Aza-dC受到影响表皮生长因子受体甲基化。在社会膜翅目昆虫中,DNA甲基化主要发生在cytosine-phosphate-guanine二核苷酸(论文认定)的外显子33,34。因为蚂蚁只生长在幼虫发育,我们测量了甲基化水平的几个其实论文认定表皮生长因子受体在prepupal阶段,这标志着幼虫的发展。两个物种,表皮生长因子受体甲基化的确是受到5-Aza-dC;每个CpG分析hypermethylated 5-Aza-dC与控制prepupae(无花果。3;电子辅助表S4)。
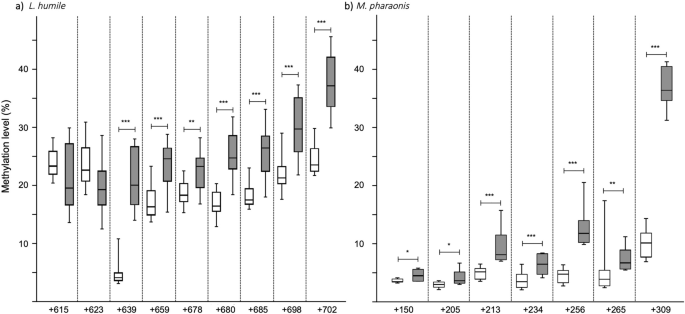
5-Aza-dC治疗的效果表皮生长因子受体甲基化。盒子的表皮生长因子受体(CpG甲基化水平一个)经由和(b)Monomorium pharaonisprepupae美联储控制解决方案(1 M蔗糖;在白色)或5-Aza-dC(灰色)当他们first-instar幼虫。从一开始的论文认定编号为每个物种最后一个外显子。每个盒子代表至少8样本数据;每个样本一个10 prepupae池。框的中线表示中位数;框的上下边缘第一和第三个四分位数,分别。须反映极端值。控制和5-Aza-dC-treated团体之间的显著差异表明:*p< 0.05,* *p< 0.01,* * *p< 0.005(学生的t测试或Mann-WhitneyU测试)。
表达水平的比较表皮生长因子受体表明这是明显在5-Aza-dC-treated prepupae(意思是褶皱变化±SD:阿根廷蚂蚁阀门= 1.18±0.78,5-Aza-dC = 9.88±1.18,p< 0.01;m . pharaonis阀门= 1.23±0.78,5-Aza-dC = 26.07±11.59,p< 0.01;无花果。4;电子辅助表S5),显示药物治疗的影响表皮生长因子受体表达式。
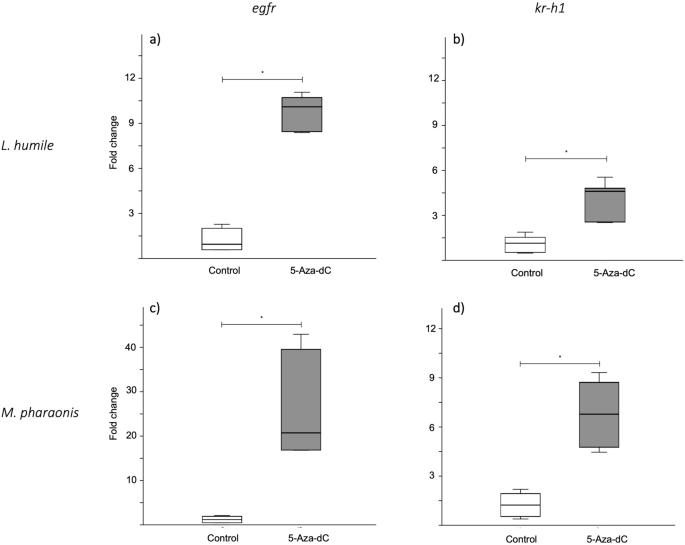
5-Aza-dC治疗的效果表皮生长因子受体和kr-h1表达式。框块相对折叠的变化(a、c)表皮生长因子受体和(b, d)kr-h1表达式为(a、b)经由和(c, d)Monomorium pharaonisprepupae美联储控制解决方案(1 M蔗糖;在白色)或5-Aza-dC(灰色)当他们first-instar幼虫。每个盒子代表数据6样品;每个样本一个10 prepupae池。框的中线表示中位数;框的上下边缘第一和第三个四分位数,分别。须反映极端值。控制和5-Aza-dC-treated团体之间的显著差异表明:*p< 0.01 (Mann-WhitneyU测试)。
表皮生长因子受体信号已知调节体型影响JH滴度11,进而调节anti-metamorphic转录因子的表达kruppel-homolog 1 (kr-h1在昆虫35,36。因此,我们相比JH-responsive基因的表达kr-h1之间的控制和5-Aza-dC-treated prepupae。至于表皮生长因子受体,我们发现5-Aza-dC-treated prepupae过表达kr-h1与控制(阿根廷蚂蚁:控制= 1.13±0.55,5-Aza-dC = 4.12±1.27,p< 0.01;m . pharaonis:控制= 1.24±0.81,5-Aza-dC = 6.79±2.01,p< 0.01;无花果。4;电子辅助表S5),这意味着JH 5-Aza-dC喂养后滴定度更高。
确认具体的表皮生长因子受体信号对工人的影响大小,我们与PD153035喂养幼虫,特定的EGFR蛋白抑制剂(EGFRi)(见方法)。正如预期的那样,对待幼虫发展成小得多的成年工人比控制幼虫(阿根廷蚂蚁唯一的t测试:p< 0.0001;无花果。5;电子辅助表S6)。
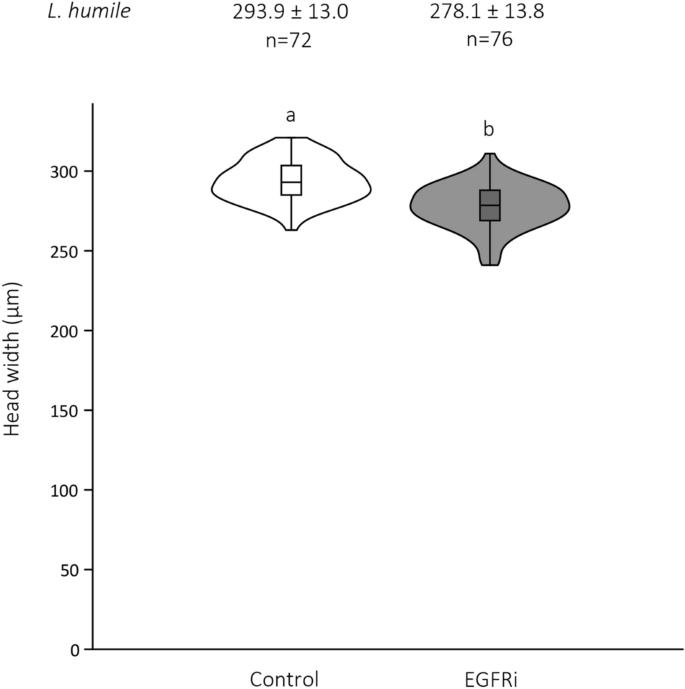
工人头上EGFRi治疗宽度的影响。小提琴头宽度的情节经由年轻而无经验的。工人的两组:幼虫美联储控制解决方案(1米蔗糖(稀释在DMSO))与幼虫美联储EGFRi(稀释在DMSO)在first-instar阶段。头宽度±SD和样本大小是上述每一个情节。框的中线表示中位数;框的上下边缘第一和第三个四分位数,分别。须反映极端值。盒子的形状是内核数据的密度分布。在小提琴上的字母表示差异在统计上有显著差异的工人头宽度(学生的t测试:p< 0.0001)。
因此,喂养幼虫的hypomethylating代理5-Aza-dC水平的提高表皮生长因子受体甲基化,表皮生长因子受体表达,kr-h1表达式,它是已知参与调节大小30.,32,35,36。始终,治疗增加工人的成人大小两个物种。
讨论
这些结果表明,工人与大小表皮生长因子受体甲基化在两个单型的蚂蚁;他们补充先前建立的关系表皮生长因子受体甲基化和工人大小与多态物种的工人30.。我们发现,工人可以药物增加大小阿根廷蚂蚁和m . pharaonis喂养幼虫hypomethylating代理,5-Aza-dC。尽管规模增长幅度相对有限,这个结果强调工人大小比观测到的塑料在自然条件下。先前的研究指出在其他蚂蚁同样的趋势37,38,39,40,41。例如,蚂蚁属大头蚁二态的:工人要么是小(未成年人)或大(专业)。当late-instar大头蚁幼虫服用JH,成年工人规模大幅增加,“超级专业”出现的时候,小说subcaste几乎没有见过39。,有趣的是,现在工人大小可塑性似乎没有充分利用在正常的生态条件。事实上,多态的工人被认为促进殖民地健身提高劳动分工6,7,8。即使在单型的蚂蚁,工人发现大小略有不同,显示不同的行为40,41,42,43,这表明即便是有限大小变异可能功能适应性。可塑性可能是工人大小限制在自然条件下,因为有工人数量和质量之间的权衡,地方小,低质量的工人创造比大便宜,高质量的员工9。这个解释符合阿根廷蚂蚁法老蚂蚁生态学。两个物种高度侵袭性,他们的竞争优势在很大程度上是由工人的丰富44,45。在一个平衡的情况下,投资于更大的工人可以把殖民地健康风险。
当阿根廷蚂蚁和m . pharaonis幼虫有5-Aza-dC,表皮生长因子受体在prepupae hypermethylated在几个论文认定。似乎违反直觉,hypomethylating代理将导致表皮生长因子受体甲基化。然而,5-Aza-dC对DNA甲基化的影响已被证明是sequence-specific46。先前的研究在社会膜翅目昆虫发现5-Aza-dC确实可以导致当地的甲基化30.,47,48,49。例如,在c . floridanus,工人幼虫美联储5-Aza-dC展出全基因组hypomethylation和基层表皮生长因子受体甲基化30.。5-Aza-dC hypermethylating效应也在植物50。这种逆关系被认为是由相声和其他表观遗传机制,如组蛋白转录后修饰51或非编码rna52。
在这两个物种,5-Aza-dC喂养诱导表皮生长因子受体超表达。这个结果符合良好的积极的关系表皮生长因子受体表达和增长53,54。工人规模下降这一事实应对EGFRi治疗证实表皮生长因子受体信号直接参与生产调节剂阿根廷蚂蚁。
我们发现5-Aza-dC治疗导致的过度kr-h1在阿根廷蚂蚁和m . pharaonis,这也符合在蚂蚁JH的促生长作用。当JH产生特定时期激素敏感性,幼虫蜕皮取代变质蜕皮,延长幼虫发育并最终促进成人体型55。这里,我们确实观察到5-Aza-dC治疗轻微增加幼虫在两个物种的持续时间(见电子补充表S3)。达到更明显发展变化,可能需要使用5-Aza-dC浓度较高或不同时间治疗。此外,串联超表达的表皮生长因子受体和kr-h1支持这个想法,表皮生长因子受体信号和JH产量呈正相关。然而,由于5-Aza-dC可能影响其他基因的甲基化,我们不能得出结论表皮生长因子受体超表达是单独负责kr-h1超表达。例如,胰岛素/胰岛素样生长因子信号通路(IIS)在膜翅目昆虫社会促进JH生产5,kr-h1超表达可能导致从IIS upregulation 5-Aza-dC。说,发生交错活化表皮生长因子受体和受体之间的生长调节途径,这意味着EGFR集由多个刺激信号提示56,57。因此,kr-h1超表达,我们观察到的最有可能导致串扰在几个调节刺激经济增长的途径,包括表皮生长因子受体信号系统。
综上所述,我们的结果表明,表皮生长因子受体甲基化和表达是始终与工人身体大小两个单型的蚂蚁属于不同亚科。必须强调两个重要的点。首先,我们检查的影响表皮生长因子受体甲基化对工人的大小,而不是在种姓的决心。的确,queen-worker种姓决心涉及许多表型变化超出身体大小。因此,我们的工作不会说种姓的决心。第二,尽管我们的研究表明之间的关系表皮生长因子受体甲基化和体型是守恒的蚂蚁,我们没有展示之间的直接因果关系表皮生长因子受体甲基化和工人的大小。值得注意的是,其他基因可能参与。进一步补充工作结合全基因组DNA甲基化分析与新兴methylome编辑技术58应该可以明确建立因果关系DNA甲基化和多样化的表型性状之间潜在的膜翅目昆虫的生态社会的成功。
方法
采样和饲养
经由收集在这个领域在接受(法国南部);Monomorium pharaonis从实验室获得殖民地饲养在哥本哈根大学生物学系、生态学、进化)。两个物种,我们建立了大量的殖民地,包含几十皇后区,成千上万的工人,育发展阶段。殖民地被保持在实验室条件下(阿根廷蚂蚁:温度= 25±1°C和相对湿度=±30%;m . pharaonis:温度= 28±1°C和相对湿度= 50%;两种:身子光/暗周期)。殖民地有糖水随意和接收新鲜的蟑螂每隔一天。
的形态学分析
量化工人大小变化,我们测量工作者头宽度(眼睛除外),一个精确的代理人员规模的蚂蚁59,60,61年。所有测量数据进行精确到0.001毫米使用MZ6立体显微镜(徕卡微系统公司,位于德国)。
药理修改DNA甲基化
幼虫被单独美联储0.5μl 10毫米5-Aza-dC(稀释蔗糖;Sigma-Aldrich A3656)或0.5μl 1 M蔗糖(控制解决方案)。然后,治疗30或30控制幼虫转移到个人实验包含300名工人,但没有皇后或窝巢。实验巢是每天检查,和新兴工人(也就是说,callow)被移除并存储在EtOH 99%后的形态学分析。因为工人是完全无菌的物种,我们确定的callow已经开发了实验幼虫。
结果显示,5-Aza-dC治疗只有显著影响成年工人大小当应用于first-instar幼虫(补充图。S1;电子辅助表S1)。因此我们关注first-instar幼虫时重复5-Aza-dC实验的物种。剩下一半的幼虫发展成成人的形态学分析。另一半是prepupal阶段收集的,flash冻结,并存储在−80°C随后的甲基化和基因表达分析。使用prepupal阶段进行分子分析源于发现5-Aza-dC改变了发展持续时间在两个物种的研究。因此,通过比较控制和5-Aza-dC幼虫在特定发展阶段(prepupae),我们确保了信号检测治疗(控制造成的vs5-Aza-dC喂养),而不是从发育率的差异。
DNA甲基化
的甲基化表皮生长因子受体用直接测量酸性亚硫酸盐测序(dBS)62年,63年,64年。我们汇集十个人从每组获得足够的生物材料。每个池被视为生物复制。
我们提取的基因组DNA使用SDS /蛋白酶K内部协议,随后呕吐phenol-chloroform /氯仿洗和乙醇/乙酸钠沉淀。用琼脂糖凝胶电泳基因组DNA碎片进行评估。量和吸光度比值测定使用一个量子位2.0荧光计(热费希尔科学)和NanoDrop 1000分光光度计(热费希尔科学),分别。基因组DNA是亚硫酸氢钠处理unmethylated胞核嘧啶转化为尿嘧啶而不影响甲基胞核嘧啶65年。亚硫酸氢盐转换进行200 - ng使用Methylamp基因组DNA的DNA样本附件(Epigentek, p - 1001)按照制造商的指示。
我们使用MethPrimer v . 2.0软件设计引物66年与一个视图放大CpG-rich地区的最后一个外显子表皮生长因子受体亚硫酸氢转换后。的序列表皮生长因子受体得到的膜翅目昆虫基因组数据库67年。PCR反应包括2×10μl多元Mastermix(试剂盒,206143),10 2μlμM正向和反向引物,5μl ddH2啊,1μl bisulphite-treated基因组DNA。循环条件如下:15分钟在95°C;35周期30年代在94°C, 90年代在54°C,和90年代在72°C;和10分钟在72°C。因为亚硫酸氢转换可以减少复杂性,因此减少PCR特异性DNA,我们提取目标PCR产品直接从琼脂糖凝胶后一夜之间进行迁移使用QIAquick凝胶萃取设备(试剂盒,28706)。
我们直接测序gel-extracted PCR产品使用3730 DNA分析仪(热费希尔科学)和BigDye终结者v . 3.1循环测序工具包(热费希尔科学,4337457)。测序反应包括2μl PCR产品6μl ddH2啊,1μl BigDye 2.1μl BigDye 5×缓冲区,和0.2μl 10μM底漆。每个生物测序使用复制技术一式三份。每个CpG甲基化水平的测量使用CodonCode对准器诉9.0.1软件(CodonCode公司)。
基因表达
我们测量表皮生长因子受体表达式使用逆转录定量实时PCR (RT-qPCR)。使用三总RNA提取试剂(热费希尔科学)。RNA分散、数量和吸光度比值评估如上所述。的相对表达表皮生长因子受体和kr-h1量化利用ΔΔCt方法吗68年和rpl32的正常化69年,70年。从膜翅目昆虫基因组数据库获得的序列65年使用NCBI引物,引物设计软件71年。
表皮生长因子受体信号通路的抑制
First-instar幼虫的阿根廷蚂蚁喂养PD153035 (EMD微孔,234491),专门一种化合物能够抑制表皮生长因子受体酪氨酸激酶活性,需要激活表皮生长因子受体途径,允许下游信号、细胞生长、细胞增殖72年。EGFRi已经成功应用在另一个蚂蚁30.。治疗幼虫有0.5μl 10μM EGFRi(稀释1 M蔗糖和DMSO)。控制幼虫有1米蔗糖(稀释在DMSO)。每组的幼虫被转移到个人实验巢穴。callow收集,他们的体型测量在一天内出现。
统计分析
我们测试数据正常和异方差性使用Shapiro-Wilk测试和列文的测试,分别。然后采用适当的统计方法(参数和非参数)。所有的统计分析使用Past4软件4.10 v73年。
数据可用性
所有生成的数据或分析在本研究中包括发表的这篇文章(和它的补充信息文件)。
引用
通过S。et al。适应性表型可塑性:共识和争议。生态发展趋势。另一个星球。10,212 - 217 (1995)。
威尔逊,e . O。昆虫社会(哈佛大学出版社,1971)。
罗宾逊,g . E。,Fahrbach, S. E. & Winston, M. L. Insect societies and the molecular biology of social behavior.BioEssays19(12),1099 - 1108 (1997)。
麦切纳,c, D。蜜蜂的世界(约翰·霍普金斯大学出版社,2007)。
电晕,M。,利R。&Wheeler, D. E. Molecular mechanisms of phenotypic plasticity in social insects.咕咕叫。当今。昆虫科学。13,则高达55 - (2016)。
Holldobler, b & Wilson e . O。蚂蚁(哈佛大学出版社,1990)。
奥斯特,g . f . & Wilson e . O。种姓制度和生态社会昆虫(普林斯顿大学出版社,1978)。
Traniello j . f .蚂蚁觅食策略。安。启Entomol。34(1),191 - 210 (1989)。
彼得斯,c & Ito f无翼和矮工作者构成的生态成功蚂蚁(膜翅目:蚁科)。Myrmecol。新闻21,117 - 130 (2015)。
La Richeliere F。et al。温暖和世界热点地区的干旱地区超个体的复杂性。Proc。r . Soc。B289年(1968),20211899 (2022)。
Nijhout h . f . &。麦凯纳k . z polyphenic发展中的胰岛素信号的不同的角色。咕咕叫。当今。昆虫科学。2558 - 64 (2018)。
小山,t &欢笑,c . k .解体机制的多样性通过营养调节昆虫体型。咕咕叫。当今。昆虫科学。25,1 - 8 (2018)。
镰仓,m . Royalactin诱发女王蜜蜂的分化。自然473年(7348),478 - 483 (2011)。
Hietakangas诉&科恩,s m .监管组织生长的营养传感。安。启麝猫。43,389 - 410 (2009)。
克鲁斯,J。,Martín, D. & Franch-Marro, X. Egfr signaling is a major regulator of ecdysone biosynthesis in the Drosophila prothoracic gland.咕咕叫。医学杂志。30.(8),1547 - 1554 (2020)。
大师,j·L。,Cobo, J. & Bellés, X. Target of rapamycin (TOR) mediates the transduction of nutritional signals into juvenile hormone production.生物。化学。284年(9),5506 - 5513 (2009)。
伯杰,s . L。,Kouzarides, T., Shiekhattar, R. & Shilatifard, A. An operational definition of epigenetics.Dev的基因。23(7),781 - 783 (2009)。
Bonasio, R。,Tu, S. & Reinberg, D. Molecular signals of epigenetic states.科学330年,612 - 616 (2010)。
Maleszka, r .表观遗传的集成环境和蜜蜂基因组信号:营养的重要的相互作用,大脑和生殖网络。表观遗传学3,188 - 192 (2008)。
Bonasio, R。et al。全基因组DNA和caste-specific methylomes的蚂蚁Camponotus floridanus和Harpegnathos saltator。咕咕叫。医学杂志。22,1755 - 1764 (2012)。
Morandin C。,Brendel, V. P., Sundström, L., Helanterä, H. & Mikheyev, A. S. Changes in gene DNA methylation and expression networks accompany caste specialization and age-related physiological changes in a social insect.摩尔。生态。28(8),1975 - 1993 (2019)。
Lyko F。et al。蜜蜂表观基因组:微分DNA甲基化的大脑在皇后区和工人。公共科学图书馆杂志。8(11),e1000506 (2010)。
Foret, S。et al。DNA甲基化动力学、代谢通量,基因拼接,在蜜蜂和替代表型。Proc。国家的。学会科学。美国109年(13)4968 - 4973 (2012)。
Kucharski, R。,Maleszka, J., Foret, S. & Maleszka, R. Nutritional control of reproductive status in honeybees via DNA methylation.科学319年(5871),1827 - 1830 (2008)。
利R。,Oxley, P. R., Keller, L. & Kronauer, D. J. C. Robust DNA methylation in the clonal raider ant brain.咕咕叫。医学杂志。26(3),391 - 395 (2016)。
Patalano, S。et al。塑料的分子签名表型在两个群居昆虫与简单的社会。Proc。国家的。学会科学。美国112年(45),13970 - 13975 (2015)。
哈里斯,k·D。,Lloyd, J. P., Domb, K., Zilberman, D. & Zemach, A. DNA methylation is maintained with high fidelity in the honey bee germline and exhibits global non-functional fluctuations during somatic development.Epigenet。染色质12(1)队(2019)。
Cardoso-Junior, c。et al。DNA甲基化的基因表达重组不是一个司机蜜蜂的年轻工人。摩尔。生态。30.(19),4804 - 4818 (2021)。
Oldroyd认为,b p & Yagound b .表观遗传学的角色,尤其是DNA甲基化,在昆虫社会进化的种姓。费罗斯。反式。r . Soc。B376年(1826),20200115 (2021)。
阿尔瓦拉多,S。,Rajakumar, R。,Abouheif, E. & Szyf, M. Epigenetic variation in the Egfr gene generates quantitative variation in a complex trait in ants.Commun Nat。66513 (2015)。
琼斯,p . a &泰勒,s m细胞分化,胞嘧啶核苷类似物和DNA甲基化。细胞20.(1),85 - 93 (1980)。
特战分队,K。,Matsuoka, Y., Funahashi, A. & Kitano, H. A comprehensive pathway map of epidermal growth factor receptor signaling.摩尔。系统。医学杂志。1(1),2005 - 2010 (2005)。
Zemach,。,McDaniel, I. E., Silva, P. & Zilberman, D. Genome-wide evolutionary analysis of eukaryotic DNA methylation.科学328年(5980),916 - 919 (2010)。
Li-Byarlay, h . DNA甲基化的功能是在社会性昆虫。前面。生态。另一个星球。457 (2016)。
Minakuchi C。,Zhou, X. & Riddiford, L. M. Krüppel homolog 1 (Kr-h1) mediates juvenile hormone action during metamorphosis of黑腹果蝇。动力机械。Dev。125年(1 - 2),91 - 105 (2008)。
Shpigler, H。et al。的转录因子Kruppel同族体1与激素介导的蜜蜂的社会组织。BMC另一个星球。医学杂志。10(1)1-13 (2010)。
Cahan, s . H。,Graves, C. J. & Brent, C. S. Intergenerational effect of juvenile hormone on offspring inPogonomyrmex收割机蚂蚁。j . Comp。杂志。B181年(8),991 - 999 (2011)。
施密特,a . M。,Linksvayer, T. A., Boomsma, J. J. & Pedersen, J. S. Queen–worker caste ratio depends on colony size in the pharaoh ant (Monomorium pharaonis)。昆虫Soc。58(2),139 - 144 (2011)。
Rajakumar, R。et al。祖先发展潜力促进平行进化的蚂蚁。科学335年,79 - 82 (2012)。
Herbers, j . m . &坎宁安,m .社会组织Leptothorax longispinosus娃。动画。Behav。31日(3),759 - 771 (1983)。
韦斯特林,j . N。,Harrington, K., Bengston, S. & Dornhaus, A. Morphological differences between extranidal and intranidal workers in the antTemnothorax rugatulus在觅食,但没有影响身体大小的距离。昆虫Soc。61年(4),367 - 369 (2014)。
Grześ,i M。,Okrutniak, M. & Grzegorzek, J. The size-dependent division of labour in monomorphic antLasius尼日尔。欧元。j .土壤。医学杂志。77年,1 - 3 (2016)。
Vele, a & Modlinger r .体型的木蚁工人会影响他们的工作部门。社会生物学66年(4),614 - 618 (2019)。
人类,k . g . &戈登,d . m .剥削和干扰入侵阿根廷蚂蚁之间的竞争,经由和本地蚂蚁。环境科学105年(3),405 - 412 (1996)。
潮湿,j·k·法老蚁的全球传播,Monomorium pharaonis(膜翅目:蚁科)。Myrmecol。新闻13,115 - 129 (2010)。
Seelan, r S。,Mukhopadhyay, P., Pisano, M. M. & Greene, R. M. Effects of 5-Aza-2′-deoxycytidine (decitabine) on gene expression.药物金属底座。牧师。50(2),193 - 207 (2018)。
做饭,N。,Parker, D. J., Tauber, E., Pannebakker, B. A. & Shuker, D. M. Validating the demethylating effects of 5-aza-2′-deoxycytidine in insects requires a whole-genome approach: (A reply to Ellers et al.).点。Nat。194年(3),432 - 438 (2019)。
博罗,我m。et al。大黄蜂的殖民地发展DNA甲基化的影响。BMC染色体组。22(1),1 - 11 (2021)。
阿马拉辛赫,h·E。,Clayton, C. I. & Mallon, E. B. Methylation and worker reproduction in the bumble-bee (Bombus terrestris)。Proc。r . Soc。B281年,20132502 (2014)。
刘,S。et al。甲基化抑制剂5-Aza-2′脱氧胞苷诱导水稻全基因组hypomethylation。大米15(1),1 - 12 (2022)。
雪松、h·伯格曼& y连接DNA甲基化和组蛋白修饰:模式和范例。Nat,启麝猫。10(5),295 - 304 (2009)。
Csankovszki G。,Nagy, A. & Jaenisch, R. Synergism of Xist RNA, DNA methylation, and histone hypoacetylation in maintaining X chromosome inactivation.j .细胞杂志。153年(4),773 - 784 (2001)。
Herbst r . s .的表皮生长因子受体生物学。Int。j . Radiat。肿瘤防治杂志。医学杂志。理论物理。59(2)S21-S26 (2004)。
Dabour, N。et al。板球体型改变系统性RNAi对胰岛素信号组件和表皮生长因子受体。Dev。增长是不同的。53(7),857 - 869 (2011)。
Nijhout h . f . &惠勒,d . e .昆虫保幼激素和生理基础的多态性。问:启杂志。57(2),109 - 133 (1982)。
奥利维拉,S。,Schiffelers, R. M., Storm, G., Henegouwen, P. M. P. & Roovers, R. C. Crosstalk between epidermal growth factor receptor-and insulin-like growth factor-1 receptor signaling: Implications for cancer therapy.咕咕叫。癌症药物靶点9(6),748 - 760 (2009)。
Berasain C。et al。表皮生长因子受体(EGFR)在肝癌相声。癌症3(2),2444 - 2461 (2011)。
刘,x。et al。编辑在哺乳动物基因组中DNA甲基化。细胞167年(1),233 - 247 (2016)。
波特,s . d .快速、准确测量蚂蚁头宽度的方法。安。Entomol。Soc。点。76年(5),866 - 867 (1983)。
彼得斯,C。鲻鱼,M。,Lin, C. C. & Billen, J. Evolution of cheaper workers in ants: A comparative study of exoskeleton thickness.医学杂志。j·林恩。Soc。121年(3),556 - 563 (2017)。
卡斯帕里,m &魏瑟·m·d·size-grain假说和种间扩展蚂蚁。功能。生态。13(4),530 - 538 (1999)。
Frommer, M。et al。基因组测序的协议,它产生一个积极的5-methylcytosine残留在单独的DNA链。Proc。国家的。学会科学。美国89年(5),1827 - 1831 (1992)。
江,M。et al。DNA甲基化的快速量化测量峰的相对高度直接bisulfite-PCR测序痕迹。实验室。Investig。90年(2),282 - 290 (2010)。
帕里什,R R。,Day, J. J. & Lubin, F. D. Direct bisulfite sequencing for examination of DNA methylation with gene and nucleotide resolution from brain tissues.咕咕叫。Protoc。>。60(1)7-24 (2012)。
Hayatsu, H。,Wataya, Y., Kai, K. & Iida, S. Reaction of sodium bisulfite with uracil, cytosine, and their derivatives.生物化学9(14),2858 - 2865 (1970)。
李,l . c &闹市区Dahiya (r . MethPrimer:为甲基化设计引物pcr。生物信息学18(11),1427 - 1431 (2002)。
Elsik, c·G。et al。膜翅目昆虫基因组数据库:整合在HymenopteraMine基因组注释。核酸Res。44d793 - 800 (2006)。
Livak, k . j . & Schmittgen t . d .相对基因表达数据的分析使用实时定量PCR和2−ΔΔCT方法。方法25(4),402 - 408 (2001)。
Gospocic, J。et al。神经肽corazonin控制蚂蚁的社会行为和种姓身份。细胞170年(4),748 - 759 (2017)。
盛,L。et al。蚂蚁的社会重组引发与长寿相关的神经胶细胞重建。科学。睡觉。6(34),eaba9869 (2020)。
你们,J。et al。Primer-BLAST:一个工具来设计有针对性的引物聚合酶链反应。BMC Bioinform。13(1),1 - 11 (2012)。
炸,d . W。et al。一个特定的表皮生长因子受体酪氨酸激酶抑制剂。科学265年(5175),1093 - 1095 (1994)。
锤子,O。,Harper, D. A. T. & Ryan, P. D. PAST: Paleonthological Statistics software package for education and data analysis.Palaeontol。电子。4(1),1 - 9 (2001)。
确认
我们感谢j。j Boomsma和r . s . Larsen提供我们m . pharaonis殖民地,j . Pearce-Duvet语言编辑服务。
资金
这项工作是支持的比利时昏聩de la任职(FRS-FNRS)(批准号T.0140.18 S.A.;FRIA奖学金。FC36257大学(Universite Libre de Bruxelles)西奥多)和罗斯福的布兰奇S.A.)(批准操作。
作者信息
作者和联系
贡献
顶替,C.G. and S.A. conceived the study; T.R. and S.A. collected biological samples on the field; T.R. carried out the experimental work and performed the data analyses; T.R. and S.A. drafted the manuscript. All authors gave final approval for the publication.
相应的作者
道德声明
相互竞争的利益
作者宣称没有利益冲突。
额外的信息
出版商的注意
施普林格自然保持中立在发表关于司法主权地图和所属机构。
权利和权限
开放获取本文是基于知识共享署名4.0国际许可,允许使用、共享、适应、分布和繁殖在任何媒介或格式,只要你给予适当的信贷原始作者(年代)和来源,提供一个链接到创作共用许可证,并指出如果变化。本文中的图片或其他第三方材料都包含在本文的创作共用许可证,除非另有说明在一个信用额度的材料。如果材料不包括在本文的创作共用许可证和用途是不允许按法定规定或超过允许的使用,您将需要获得直接从版权所有者的许可。查看本许可证的副本,访问http://creativecommons.org/licenses/by/4.0/。
关于这篇文章

引用这篇文章
狐狸,T。,Gueydan, C. & Aron, S. DNA methylation and expression of the表皮生长因子受体基因与工人在单型的蚂蚁大小有关。Sci代表1221228 (2022)。https://doi.org/10.1038/s41598 - 022 - 25675 - 4
收到了:
接受:
发表:
DOI:https://doi.org/10.1038/s41598 - 022 - 25675 - 4
