















«上一页«上一页下一个»

达尔文的“奥秘中的奥秘”
"...这些形式可能仍然只是…品种;但是,我们只要假定,要把这些形式转化为物种,改造的步骤就会更多,或者在数量上更大。因此物种繁殖(达尔文1859,第120页)。
进化生物学中大多数话题的讨论都是从达尔文开始的。事实上,《物种起源》(1859)继续影响着现代进化生物学。达尔文通过自然选择作为种群内部变化的一个非常渐进的机制,并假设新物种可能是这个过程的产物,但需要更长的时间。物种形成的最终过程自然选择在《物种起源》出版近20年前,达尔文在他的个人笔记本上画了一张草图(图1)。在这里,他提出了一个模型,即血统是通过在相对较长的时间内进化不同的特征来从祖先形成的。达尔文指出,物种可以通过一个物种分裂成两个物种的进化来形成,或者通过一个种群从现有的祖先分化成一个新物种来形成。达尔文对进化的洞见是杰出的,尤其是在没有遗传因素的情况下。事实上,关于遗传和通过突变引入新的遗传物质的想法在达尔文提出进化论之后很久才出现。
现代综合
进化研究的一个重大转折点发生在20世纪30年代,当时Fisher、Haldane、Wright、Dobzhansky等人开发了数学种群遗传模型,以阐明进化变化的遗传机制(Mayr & Provine 1998)。遗传学与自然选择模型的结合极大地阐明并加强了达尔文关于进化的观点——这是通过突变和重组将新的变异引入种群的缺失机制。的确,多亏了现代综合,目前进化生物学的许多研究都与遗传学紧密相关,目前研究物种形成的方法也不例外。正如下面所讨论的,现代综合理论不仅在种群进化研究方面取得了进展,而且在物种定义方式和新物种形成方式方面也发生了变化。
繁殖障碍。
根据常用的“生物物种概念”(Mayr 1942),新物种的形成涉及在交配前(合子前障碍)或交配后不久(合子后障碍)繁殖障碍的进化,以产生可存活的后代。因此,当来自不同种群的个体不再将彼此视为潜在的配偶,或者交配机会因栖息地使用或繁殖时间表的差异而受到限制时,新物种就形成了。在某些情况下,这些合子前的隔离机制不能防止来自不同群体的个体之间的杂交繁殖。在这些情况下,可能会形成可存活的杂交体,或者一次成功的交配尝试可能以失败告终,原因要么是产生了不能存活的合子,要么是产生了不育的、不能繁殖的后代。这些不同的合子前和合子后障碍对物种形成生物学家来说非常重要,因为它们决定了繁殖隔离的种群彼此之间的距离,这表明了种群在通常连续的物种形成过程中走了多远。例如,生殖隔离在物种形成的早期阶段是弱的,但在物种形成的后期阶段会转变为强的或完全的(图2)。许多类型的隔离机制中的一种或多种可能在物种沿连续统的进化中发挥作用(图2)。但是遗传交换的生殖障碍是如何以及为什么会进化?
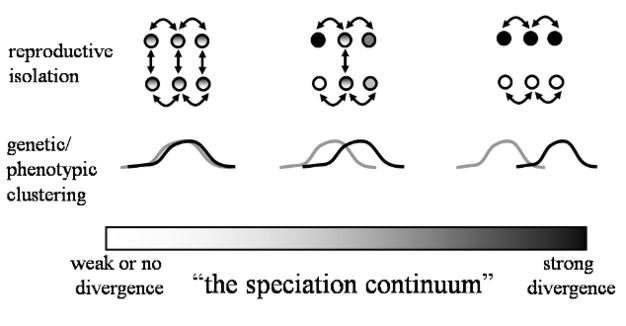 图2:物种形成过程中发散的连续性质的示意图,沿物种形成连续统描绘了三个任意点。
许多类型的分化在数量上有所不同,分化的大小代表了物种形成的程度。两个箭头代表个体之间的交配。
©2012自然教育经Nosil允许修改和转载et al。2009.版权所有。
图2:物种形成过程中发散的连续性质的示意图,沿物种形成连续统描绘了三个任意点。
许多类型的分化在数量上有所不同,分化的大小代表了物种形成的程度。两个箭头代表个体之间的交配。
©2012自然教育经Nosil允许修改和转载et al。2009.版权所有。
![]()
地理在物种形成中的作用。
物种生物学家之间争论的一个主要领域是物种形成的地理环境(图3)。恩斯特·迈尔(Ernst Mayr)强调捍卫了他的观点,即物种形成最有可能发生在种群在地理上相互隔离的时候,这样孤立种群内的进化将导致它们之间的足够差异,从而导致物种形成将是最终的结果。”…作为稳定的遗传分化的副产品,隔离机制的进化是不可避免的(Mayr 1963,第581页)。这里的中心思想是,当人口在地理上分离时,他们会在外观和基因上彼此产生分歧。这些变化可能通过自然选择或随机机会(即遗传漂变)发生,在这两种情况下都会导致生殖隔离。这种地理上孤立种群的物种形成观点——被称为异域物种形成——仍然被物种生物学家广泛认为在生物多样性的进化中发挥着重要作用(例如,Price 2007)。然而,物种形成也可能发生在地理上不孤立的重叠种群中(即同域物种形成,Via 2001)。这里的问题是,生活在同一地区并交换基因的人群是如何彼此分化的?许多生物学家认为这将是极其困难的(Coyne & Orr 2004),但有一些令人信服的例子,不同栖息地的种群受到不同自然选择模式的影响(即,分歧选择),并克服基因流,分化为不同的物种。这可能发生,例如,如果昆虫适应了生活在同一地理区域内的不同植物(Feder等.1988)。随着同域物种形成的争议逐渐减少,看看会有多少新的例子出现将是一件有趣的事情。面对基因流,物种形成的另一种情况是“跨物种形成”,尽管其水平低于同域物种形成。Parapatric speciation指的是地理位置接近的种群,通常是相邻但不重叠的范围。在这里,每个种群中都有一小部分彼此有实际接触,因此被认为是同域性的,而大多数个体居住在足够远的地方,彼此之间很少频繁相遇(图3)等.2008年)和手杖昆虫(Nosil等.(Coyne & Orr 2004),但这种现象比异域或同域物种形成(allopatric speciation)受到的关注要少。









