


 图1:Pre-mRNA拼接。
图1:Pre-mRNA拼接。

















«上一页下一个»
对于大多数真核生物的基因(和一些原核的),最初的核糖核酸从基因的转录DNA模板之前必须处理它就变成了一个成熟信使核糖核酸(信使核糖核酸),可以直接的合成蛋白质。这个处理的步骤之一,被称为RNA拼接,包括删除或某些序列称为的“拼接”序列,干预或内含子。最后的mRNA因此由剩余的序列,称为外显子,通过拼接连接到另一个过程。RNA拼接最初发现于1970年代,推翻了多年的思想领域的基因表达式。
早期的研究在细菌中
基因调控首次研究了相对简单的细菌中最彻底的系统。大多数细菌RNA转录并不进行拼接;这些记录是共线的,直接与DNA编码。换句话说,有一一对应的基地之间的基因和基因的mRNA转录(除了5′和3′非编码区域)。然而,在1977年,几组研究人员与腺病毒的感染和复制在哺乳动物细胞获得一些令人吃惊的结果。这些科学家发现了一系列的RNA分子,称为“马赛克”,每一个都包含从非邻接网站在病毒序列基因组(大山大山等,1977;周润发et al。,1977)。这些马赛克在病毒感染中被发现。早期感染的研究显示长期主要包含所有的RNA转录,晚期RNA序列,以及被称为干预序列(内含子)。
随后adenoviral发现,内含子中发现了许多其他病毒和真核基因,包括血红蛋白和免疫球蛋白(达内尔,1978)。拼接的RNA转录中观察到几个在体外系统来自真核细胞,包括去除内含子转移核糖核酸在酵母无细胞提取物(克纳普等,1978)。这些观察固化大型初始的假设拼接成绩单了,事实上,产生成熟的信使rna。其他假设提出DNA模板以某种方式循环或假定一个允许的二级结构转录从非邻接区域(达内尔,1978)。
如何剪接发生
", "true", "All rights reserved.", '900', '682', '//www.scienovate.com');">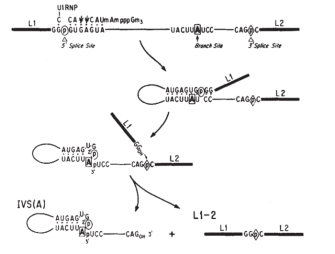

", "true", "All rights reserved.", '900', '682', '//www.scienovate.com');">拼接pre-mRNA分子发生在几个步骤所催化的小型核核糖核蛋白(snRNPs)。后U1 snRNP结合5′拼接网站,5′末端插入碱基对的下游分支序列,形成一个套索。外显子3′末端的切割和加入到分支站点的3′末端羟基(OH)小组攻击的磷酸二酯键的外显子3′拼接的网站。因此,外显子(L1和L2)共价结合,并包含内含子释放的套索。
©1985自然出版集团Konarska, M . M。et al。描述分支站点的套索rna拼接产生的信使rna前体。自然
313年,556 (1985)。保留所有权利。
", '900','//www.scienovate.com', "In the first step of RNA splicing, the pre-mRNA is depicted as a line. From left (the 5′ end) to right (the 3′ end), the pre-mRNA is labeled as follows: a solid section is labeled L1, which stands for leader exon 1. L1 is followed by the sequence at the 5′ end of the intron: GGpGUGAGUA. The second G in this sequence is bound to a small nuclear ribonucleoprotein labeled U1RNP. The p is a phosphodiester bond labeled 5′ splice site. Following an unlabeled segment is the branch site sequence: UACUUAUCC. The second A in this sequence is labeled branch site. Following a second unlabeled segment is the 3′ end of the intron: CAGpC. The p is a phosphodiester bond labeled 3′ splice site. Lastly, the pre-mRNA has a solid section labeled L2, which stands for leader exon 2. An arrow leads to the second step. The sequence at the 5′ end of the intron base pairs with the conserved branch point region downstream to form a looped structure called a lariat. In the third step, the 3′ end of the exon L1 is cut, and a hydroxyl (OH) group at the 3′ end of the exon attacks the phosphodiester bond at the 3′ splice site. In the final step, the L1 and L2 exons are covalently bound, and the resulting lariat containing the intron is released.")" class="inlineLinks">细节图
剪接发生在几个步骤,由小核核糖核蛋白(催化snRNPs“snurps”),通常明显。首先,pre-mRNA裂解的5′末端基因内区下面的附件snRNP称为U1的互补在内含子序列。然后切端连接到守恒的分枝点区域下游通过配对鸟嘌呤和腺嘌呤核苷酸5′末端分叉点,分别称为形成毛圈结构套索(图1)。鸟嘌呤和腺嘌呤碱基的结合通过化学反应称为发生酯交换中,碳原子上的羟基(OH)组的腺嘌呤鸟嘌呤核苷酸的“攻击”债券拼接站点。鸟嘌呤的残渣因此从RNA链裂解,形成一个新的债券与腺嘌呤。
接下来,snRNPs U2和U4 / U6似乎有助于定位5′末端和分支点的距离。参与U5, 3′末端的基因内区进入邻近,削减,并加入5′末端。这一步发生酯交换;在这种情况下,一个羟基的3′末端外显子攻击磷酸二酯债券3′接头地点。临近的外显子是共价结合,生成的套索释放与U2, U5, U6绑定到它。
除了共识序列拼接的网站,真核基因长内含子也包含其实拼接增强剂(es)。这些序列,帮助位置拼接装置,在基因的外显子和结合蛋白,帮助招募拼接机械到正确的网站。外显子之间最剪接发生在一个RNA转录,但偶尔trans-splicing发生,外显子在不同pre-mRNAs绑定在一起。
拼接过程发生在细胞的机器称为剪接体,在发现snRNPs加上额外的蛋白质。剪接体的主要品种是最丰富的细胞结构,最近,一个二级类型的剪接体已被确认处理一个小类别的内含子。这些基因内区被称为U12-type内含子,因为他们的行动取决于snRNP叫U12(上述常见的内含子被称为U2-type内含子)。的角色U12-type内含子还没有定义,但他们的持久性进化和保护之间同源基因的大相径庭物种提出了一个重要的功能基础(帕特尔&施泰茨,2003)。
Self-Splicing和可变剪接
一些RNA分子有能力自己拼接;最初发现这self-splicing原生动物的能力四膜虫thermophila认可了1989年的诺贝尔奖。的self-splicing内含子中发现t . thermophila现在被称为组我内含子;这个类还包括其他原生动物核糖体核糖核酸基因,一些真菌线粒体基因,一些噬菌体基因。组我内含子与9个循环和折叠成一个复杂的二级结构采用酯交换反应如上所述。另一方面,第二组self-splicing内含子中发现线粒体基因和切除的一种机制,熊相似pre-mRNA拼接,包括套索的生产。出于这个原因,有人提议也许pre-mRNA内含子和剪接机制发展集团第二内含子。
早期在拼接过程中研究,另一个令人惊讶的发现是,具体地说,研究人员注意到,不仅是pre-mRNA被内含子,需要切除,而且在单个pre-mRNA拼接的替代模式分子可以产生不同的功能mrna(图2;大山大山等。1977)。第一个例子的可变剪接于1977年腺病毒中定义,证明了一个pre-mRNA分子可以在不同的拼接连接导致各种成熟的信使rna分子,每个包含不同的外显子的组合。
不久之后,发现了可变剪接发生在细胞的基因,与第一个示例中标识IgM基因、免疫球蛋白超家族的一个成员(早期et al。,1980)。基因的另一个例子,一个令人印象深刻的是可变剪接模式Dscam基因果蝇,参与指导胚胎神经目标在果蝇的神经系统的形成。考试的Dscam序列揭示了大量的内含子,微分拼接,理论上,创建一个惊人的38000种不同的mrna。这能力创造如此多的mrna可能提供必要的多样性形成一个复杂的结构,如神经系统(笨蛋et al。,2000)。事实上,存在多个在单个基因mRNA转录本可能占一些生物体的复杂性,如人类基因相对较少(约20000)。例如,从王et al。(2008)表明,超过90%的人类基因或者拼接。
内含子的过去和未来
内含子的存在和微分拼接有助于解释如何创建新基因在进化过程中。剪接基因更“模块化”,允许新外显子的组合在进化过程中被创建。此外,旧可以插入新的外显子内含子,创造新的蛋白质在不影响函数古老的基因。
我们的知识的RNA拼接很新。尽管如此,因为几乎所有真核生物有内含子和共享机制的RNA拼接,拼接本身必须是非常古老的。“intron-early”理论的支持者认为所有生物(包括原核生物)一次基因中内含子但后来失去了这些元素,而“intron-late”的支持者认为,内含子,真核生物的限制表明最近的介绍(Roy &吉尔伯特,2006)。没有明显的模式在真核生物有内含子,这让研究人员很难预测内含子是如何通过进化获得或丢失。然而,清楚的是,内含子剪接在进化过程中都扮演了重要的角色,和科学家才刚刚开始发现这个角色的本质。
引用和推荐阅读
大山大山,s M。等。拼接部分腺病毒5 '末端的mRNA 2晚。美国国家科学院院刊》上74年3171 - 3175 (1977)
周润发,l . T。等。一个了不起的顺序安排在腺病毒5′2信使RNA。细胞121 - 8 (1977)
达内尔,j·E。小的影响RNA-RNA拼接在真核细胞的进化。科学202年1257 - 1260 (1978)doi: 10.1126 / science.364651
早期,P。等。两个信使RNA可以产生从一个免疫球蛋白可变RNA链的处理途径。细胞20.313 - 319 (1980)
克纳普,G。,等。干预序列的转录和处理酵母tRNA基因。细胞14221 - 236 (1978)
Konarska, M . M。等。描述分支站点的套索rna拼接产生的信使rna前体。自然313年552 - 557 (1984)doi: 10.1038 / 313552 a0 (文章链接)
帕特尔,A。&施泰茨,j . a .拼接双:从第二个剪接体。自然4960 - 970 (2003)doi: 10.1038 / nrm1259 (文章链接)
皮尔斯,b。遗传学:概念的方法,第二版。(纽约,弗里曼,2000)
罗伊,s W。&吉尔伯特,w的进化spliceosomal内含子:模式、拼图、和进步。自然遗传学评论7211 - 221 (2006)doi: 10.1038 / nrg1807 (文章链接)
笨蛋,D。等。果蝇Dscam是一个轴突引导受体表现出非凡的分子多样性。细胞101年671 - 684 (2000)










