
文章|
-
-
-
文章|
 肺泡内毛细血管细胞型特化
肺泡内毛细血管细胞型特化肺泡是慢性疾病和病毒诱导的肺损伤的部位,对肺泡血管的单细胞分析揭示了两种混合的内皮细胞类型,具有专门的气体交换和干细胞功能。
- 阿斯特丽德Gillich
- ,张的粉丝
- &罗斯·j·梅泽尔
-
文章|
 器官发生和肿瘤发生的适应性血流动力学内皮细胞
器官发生和肿瘤发生的适应性血流动力学内皮细胞ETV2在成人内皮细胞中的短暂再激活重新编程这些细胞成为适应性强的血管生成内皮细胞,在三维矩阵中自组装成血管网络,可以运输血液和生理上树形化类器官和脱细胞组织。
- 拨立柴Palikuqi
- ,阮杜休
- &萨因Rafii
-
-
信|
 β1整合素的机械感应诱导肝脏生长和存活的血管分泌信号
β1整合素的机械感应诱导肝脏生长和存活的血管分泌信号在小鼠和人肝脏模型中,血管灌注和机械拉伸释放内皮细胞的血管分泌信号,导致肝细胞存活和肝脏生长。
- 琳达·洛伦兹
- ,詹妮弗Axnick
- &埃克哈特·拉默特
-
文章|
 谷氨酰胺合成酶在谷氨酰胺合成外血管生成中的作用
谷氨酰胺合成酶在谷氨酰胺合成外血管生成中的作用谷氨酰胺合成酶在血管生成过程中通过自噬细胞化和RHOJ信号的调节在内皮细胞迁移中活跃。
- 家伙Eelen
- ,夏洛特杜波依斯
- &彼得Carmeliet
-
文章|
 构建冠状动脉的早期祖细胞的单细胞分析
构建冠状动脉的早期祖细胞的单细胞分析在发育过程中,新的动脉可以从已经存在的静脉中产生;所涉及的细胞命运开关在小鼠胚胎心脏血流开始之前逐渐发生。
- Tianying苏
- ,杰夫·斯坦利
- &克里斯蒂红马
-
信|
 Cloche是一种bHLH-PAS转录因子,可驱动血液血管规范
Cloche是一种bHLH-PAS转录因子,可驱动血液血管规范的斑马鱼钟形大多数内皮细胞和造血细胞的形成都需要基因,但一直难以分离;这项研究揭示了钟形编码含有pas结构域的bHLH转录因子,哺乳动物同源物可以部分挽救钟形突变体,表明可能在哺乳动物中起保守作用。
- 斯文Reischauer
- ,奥利弗·a·斯通
- &Didier Y. R. Stainier
-
信|
 造血干细胞血管生态位的年龄依赖性调节
造血干细胞血管生态位的年龄依赖性调节骨内皮细胞中的Notch信号通路诱导环境中的毛细血管和间充质干细胞发生变化,以支持造血干细胞扩增。
- Anjali P. Kusumbe
- ,Saravana K. Ramasamy
- &拉尔夫·h·亚当斯
-
-
文章|
 脂肪酸碳是内皮细胞合成dNTP所必需的
脂肪酸碳是内皮细胞合成dNTP所必需的本研究确定了脂肪酸氧化(FAO)在血管生成过程中在内皮细胞中的关键作用,并揭示了脂肪酸衍生的碳用于血管生成新创因此,FAO通过增加内皮细胞增殖来刺激血管发芽。
- 桑德拉Schoors
- ,Ulrike Bruning
- &彼得Carmeliet
-
-
信|
 内皮Notch活性促进骨中的血管生成和成骨
内皮Notch活性促进骨中的血管生成和成骨研究表明,骨骼中的血管生长需要Notch信号,并涉及一种不涉及内皮芽的特殊形式的血管生成。
- Saravana K. Ramasamy
- ,Anjali P. Kusumbe
- &拉尔夫·h·亚当斯
-
文章|
 骨中特定血管亚型的血管生成和成骨耦合
骨中特定血管亚型的血管生成和成骨耦合骨稳态和修复随年龄增长而下降,而调节骨生长和血管形成之间关系的机制尚不清楚;这项小鼠研究确定了促进新骨形成的内皮细胞,这是一种可以通过高CD31和高Emcn表达来识别的小微血管亚型。
- Anjali P. Kusumbe
- ,Saravana K. Ramasamy
- &拉尔夫·h·亚当斯
-
信|
 在出生后的关键时期,神经活动紊乱会破坏脑血管生成
在出生后的关键时期,神经活动紊乱会破坏脑血管生成在小鼠体内,在出生后发育的关键时期,由重复的声音、胡须弯曲、运动活动或癫痫引起的慢性刺激,会导致大脑微血管密度永久性降低,这种影响会损害神经元的氧气输送,但可以通过阻断一氧化氮信号来防止。
- 克里斯蒂娜Whiteus
- ,卡塔琳娜州Freitas
- &Jaime Grutzendler
-
信|
 aibp介导的胆固醇外排控制血管生成
aibp介导的胆固醇外排控制血管生成基因表达研究用于阐明胆固醇调节与血管生成之间的关系:载脂蛋白A-1结合蛋白(AIBP)被发现增强了胆固醇从内皮细胞向高密度脂蛋白的内流,由此导致的胆固醇消耗改变了血管中的膜脂顺序,导致血管内皮生长因子受体信号通路降低。
- Longhou方
- ,Soo-Ho崔
- &尤里·米勒
-
文章|
 线性泛素特异性去泛素酶调节血管生成
线性泛素特异性去泛素酶调节血管生成这项研究确定了一种特异性识别和切割线性泛素链的去泛素酶(DUB),涉及Wnt信号传导和血管生成中的线性(去)泛素化;胶胶的突变导致小鼠血管生成缺陷,结构和生化分析表明,胶胶编码线性泛素特异性DUB。
- 埃琳娜Rivkin
- ,斯蒂芬妮·m·阿尔梅达
- &Sabine P. Cordes
-
信|
 一种直接和依赖黑视素的胎儿光反应调节小鼠的眼睛发育
一种直接和依赖黑视素的胎儿光反应调节小鼠的眼睛发育在视网膜血管发育过程中,玻体血管同时退行,视网膜血管形成;研究表明,发育中的脉管系统的退化是光依赖的,并通过光感受器黑视素起作用。
- 苏饶。
- ,克里斯蒂娜春
- &理查德·a·朗
-
文章|
 心脏血管生成失衡导致围生期心肌病
心脏血管生成失衡导致围生期心肌病来自小鼠和人类的证据表明,围生期心肌病(PPCM)是一种由妊娠围生期过度抗血管生成信号引起的血管疾病,先兆子痫和多胎妊娠是发生PPCM的重要危险因素。
- 伊恩·彭定康
- ,至今为止Rana
- &Zoltan Arany
-
-
-
-
信|
 肝再生需要窦内皮诱导的血管分泌信号
肝再生需要窦内皮诱导的血管分泌信号这些作者描述了血管内皮细胞在手术切除后维持肝脏再生的分子途径。在特定的肝内皮细胞亚群中,血管内皮生长因子- a受体-2的激活导致内皮特异性转录因子的上调Id1,进而诱导内皮细胞分泌Wnt2和肝细胞生长因子,引发肝细胞增殖。
- Bi-Sen叮
- ,丹尼尔·j·诺兰
- &萨因Rafii
-
信|
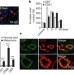 氧化应激通过新的内源性配体激活TLR2来诱导血管生成
氧化应激通过新的内源性配体激活TLR2来诱导血管生成这里表明,脂质氧化的最终产物- ω-(2-羧乙基)吡咯和其他相关吡咯-在炎症和伤口愈合过程中产生,并在小鼠的衰老组织和小鼠和人类黑素瘤的高度血管化肿瘤中以高水平积累。这些羧基吡咯被内皮细胞上的toll样受体2识别,引发了一系列导致新血管生长的事件。
- 肖夏Z.西
- ,尼古拉·马利宁
- &Tatiana V. Byzova
-
信|
 Ephrin-B2控制vegf诱导的血管生成和淋巴管生成
Ephrin-B2控制vegf诱导的血管生成和淋巴管生成已知蛋白ephrin-B2在血管生成(新血管的生长)过程中上调,但其确切功能尚不清楚。这里表明,信号通过ephrin-B2控制血管发芽。在机制上,ephrin-B2似乎部分通过调节血管内皮生长因子受体(VEGFRs)的内化发挥作用。结果表明,阻断ephrin-B2信号通路可能是阻断VEGFR功能干扰肿瘤血管生成的替代方法。
- Yingdi王
- ,Masanori Nakayama
- &拉尔夫·h·亚当斯
-
信|
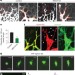 Ephrin-B2调节VEGFR2在发育和肿瘤血管生成中的功能
Ephrin-B2调节VEGFR2在发育和肿瘤血管生成中的功能已知蛋白ephrin-B2在血管生成(新血管的生长)过程中上调,但其确切功能尚不清楚。这里表明,信号通过ephrin-B2控制血管发芽。在机制上,ephrin-B2似乎部分通过调节血管内皮生长因子受体(VEGFRs)的内化发挥作用。结果表明,阻断ephrin-B2信号通路可能是阻断VEGFR功能干扰肿瘤血管生成的替代方法。
- Suphansa Sawamiphak
- ,Sascha塞德尔
- &帕罗Acker-Palmer
-
信|
 血管生成过程中microrna介导的血流动力学和Vegf信号的整合
血管生成过程中microrna介导的血流动力学和Vegf信号的整合在胚胎发育期间,血管根据血液流动而重塑。在这里,我们描述了一种遗传途径,通过这种机械感觉刺激与早期发育信号相结合来重塑斑马鱼的主动脉弓血管。发现流动诱导转录因子klf2a需要诱导内皮特异性microRNA的表达,通过生长因子Vegf激活信号。
- 普Nicoli
- ,克莱夫·史坦利
- &内森·d·劳森
-
文章|
 冠状动脉是通过静脉细胞的发育重编程形成的
冠状动脉是通过静脉细胞的发育重编程形成的目前流行的模型认为,心脏发育中的冠状动脉是由起源于心外膜的祖细胞形成的。然而,在这里发现,这些动脉是由将循环血液返回胚胎心脏的大静脉的新生血管芽产生的。因此,一些分化的静脉细胞保留发育可塑性,并对局部信号作出反应,转化为冠状动脉、毛细血管和静脉。
- 克里斯蒂红马
- ,Hiroo上野
- &马克·a·克拉斯诺
 通过淋巴转分化产生特化血管
通过淋巴转分化产生特化血管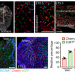 动脉化需要及时抑制细胞生长
动脉化需要及时抑制细胞生长 红髓祖细胞为血管提供内皮细胞
红髓祖细胞为血管提供内皮细胞 FOXO1在血管内皮中的代谢活性与生长状态密切相关
FOXO1在血管内皮中的代谢活性与生长状态密切相关 血管结构与生理力的压电集成
血管结构与生理力的压电集成 血管生态位和VEGF-Nrp1环调控皮肤肿瘤的起始和干性
血管生态位和VEGF-Nrp1环调控皮肤肿瘤的起始和干性 髓系细胞中非典型Wnt-Flt1通路对血管生成的调控
髓系细胞中非典型Wnt-Flt1通路对血管生成的调控 血管生成的分子机制及临床应用
血管生成的分子机制及临床应用