
文章
|开放获取
特色
-
-
文章
|开放获取 Evolvability-enhancing突变健身景观的RNA和蛋白质
Evolvability-enhancing突变健身景观的RNA和蛋白质可进化性本身是否可以适应达尔文进化的产物是一个争论不休的问题。这个研究提出适应性景观港突变,提高发展的极端分子和帮助这些分子进化种群高适应性。
- 安德烈亚斯•瓦格纳
-
文章
|开放获取 癌症基因组容忍有害编码通过体细胞拷贝数变异放大的野生型区域
癌症基因组容忍有害编码通过体细胞拷贝数变异放大的野生型区域大多数癌细胞的突变积累是有害的,目前还不清楚如何容忍这些改变。在这里,作者提出,拷贝数放大可能增加有害突变的宽容,和分析功能,可以确定潜在的选择过程。
- 法比奥Alfieri
- ,朱里奥Caravagna
- &马丁·h·奇科夫
-
文章
|开放获取 秘密的识别两个蛋白质折叠之间的进化途径
秘密的识别两个蛋白质折叠之间的进化途径蛋白质二级structures-α-helicesβ-sheets-are一般认为是固定在进化的历史。通过利用序列信息和敏感的统计技术,这项工作提出了在天然dna结合蛋白质二级结构响应转向逐步突变。
- Devlina的查克推瓦蒂
- ,Shwetha Sreenivasan
- &劳伦·l·波特
-
文章
|开放获取 定量评估可以在不完全信息下稳定间接互惠
定量评估可以在不完全信息下稳定间接互惠间接互惠描述如何合作出现在一个社区成员值他们的声誉。在这里,作者表明,细致的评估观测可以减轻当社区成员的意见分歧和错误并不同步。
- 劳拉·施密德
- ,Farbod Ekbatani
- &Krishnendu Chatterjee
-
文章
|开放获取 BdLT-Seq作为条形码decay-based方法解开lineage-linked转录组可塑性
BdLT-Seq作为条形码decay-based方法解开lineage-linked转录组可塑性细胞的可塑性是一个核心生物过程;然而,观察非基因遗传的多样性和由此产生的表型输出,是具有挑战性的。在这里作者开发基于葡萄糖跟踪技术可以用来揭示lineage-linked转录组可塑性。
- Yelyzaveta Shlyakhtina
- ,比安卡Bloechl
- &Maximiliano m .门户
-
文章
|开放获取 资源共享是充分的劳动分工的出现
资源共享是充分的劳动分工的出现分工,小组的成员专门研究在不同的任务,是许多社会有机体的核心功能。使用一个理论模型,作者证明劳动分工可以自发地出现在一群完全相同的个体。
- 简·j·罗斯
- ,Thijs简森
- &弗朗茨·j·韦斯兴起
-
文章
|开放获取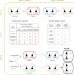 声誉效应驱动的共同进化和社会有益的合作
声誉效应驱动的共同进化和社会有益的合作奖励可以激励人们合作,但奖励行为的演变本身就是知之甚少。在这里,一个博弈论的分析表明,声誉效应促进合作和社会奖励政策的同步进化。
- Saptarshi朋友
- &基督教Hilbe
-
文章
|开放获取 母性和女性更强的免疫反应
母性和女性更强的免疫反应女性通常山感染比男性更强的免疫反应,导致更高的自身免疫性疾病的影响。在这里,作者表明,病原体传播从孕期母婴驱动器的共同进化的防御无害的病原体在女性。
- 埃文·米切尔
- ,安德里亚·l·格雷厄姆
- &Geoff野生
-
评论
|开放获取 异配生殖解释了为什么通过额外的交配雄性获益更多
异配生殖解释了为什么通过额外的交配雄性获益更多为什么男性通常比女性更激烈争夺交配机会,这与性别差异在配子大小如何?一项新的研究提供了一个正式的进化联系配子大小二态性和贝特曼梯度,它描述了多少个人的性受益于额外的交配。
- 乔纳森·m·亨肖
- ,亚当·g·琼斯
- &卢卡斯Scharer
-
文章
|开放获取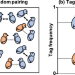 多个社会遇到可以消除牧杖的悖论和稳定遗传亲缘识别
多个社会遇到可以消除牧杖的悖论和稳定遗传亲缘识别牧杖的悖论表明,遗传亲缘识别不会进化稳定。在这里,作者表明,允许多个社会遇到之前每个社会互动可以消除牧杖悖论和稳定遗传亲缘识别。
- 托马斯·w·斯科特
- ,艾伦Grafen
- &斯图亚特·a .西
-
文章
|开放获取 贝特曼梯度从第一原则
贝特曼梯度从第一原则1948年,贝特曼宣称性选择的性别差异是由配子数。Lehtonen礼物数学模型广泛验证这一具有争议性的要求,同时指出选择在某些条件下可以扭转。
- Jussi Lehtonen
-
文章
|开放获取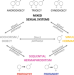 开关、稳定和逆转在鱼的性系统的进化历史
开关、稳定和逆转在鱼的性系统的进化历史鱼有一个多样性的性系统。解放军等人分析了转换在鱼,这些系统支持同时雌雄同体性不能直接从单独的性别,但需要发展顺序雌雄同体性作为中间步骤。
- 苏珊娜解放军
- ,奇亚拉Benvenuto
- &弗朗西斯皮斐尔
-
文章
|开放获取 异常相似特征的进化过渡到出生在phrynosomatid蜥蜴
异常相似特征的进化过渡到出生在phrynosomatid蜥蜴有五个独立的过渡从产卵phrynosomatid活产的蜥蜴。这里,Dominguez-Guerrero等人确定平行的生理变化,生活史和行为描述这些转换活产。
- 扫罗f . Dominguez-Guerrero
- ,Fausto r . Mendez-de la Cruz
- &玛莎·m·穆尼奥斯
-
文章
|开放获取 利基市场扩张和自适应全球辐射发散的乌鸦,乌鸦
利基市场扩张和自适应全球辐射发散的乌鸦,乌鸦特征,促进新环境的适应性反应,可能促进全球辐射。在这里,作者描述乌鸦的多样化动力学,发现他们的全球辐射的同时,高水平的表型和气候生态进化。
- 琼Garcia-Porta
- ,丹尼尔·索尔
- &卡洛斯·a·波特罗
-
文章
|开放获取 之间的权衡减少突变积累和增加组织分化决定了组织的承诺
之间的权衡减少突变积累和增加组织分化决定了组织的承诺接受更多的观察,组织干细胞分裂更容易罹患癌症提供了一个悖论,因为这些组织应该有更多的机会累积致癌突变。在这里,作者提出一个解决悖论通过展示层次组织可以通过平衡保持低癌症发病率突变积累和分化细胞的承诺。
- Marton得墨忒耳
- ,Imre Derenyi
- &Gergely j . Szollősi
-
文章
|开放获取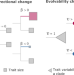 一般统计模型表明,宏观进化论模式和流程符合达尔文的渐进主义
一般统计模型表明,宏观进化论模式和流程符合达尔文的渐进主义“大进化达尔文和后来的理论家所带来的困难,因为物种经常突然改变,或经历长时间的停滞,这两个计数器增量变化或渐进主义理论。在这里,作者提出一个宏观进化论统计模型适应这种不均匀进化的景观,甚至表明突然宏观进化论更改兼容渐进microevolutionary过程。”
- 马克·佩格尔
- ,席亚拉,马路
- &安德鲁·米德
-
文章
|开放获取 合作交流与内存约束的交互
合作交流与内存约束的交互在许多情况下的互惠、个人轮流合作。在这里,作者分析了平衡和交替的动态游戏,特别是描述所有与一轮记忆策略,保持合作。
- 彼得s公园
- ,马丁·a·诺瓦克
- &基督教Hilbe
-
文章
|开放获取 选择力会随着年龄的增长,因为老龄化的发展,而不是反之亦然
选择力会随着年龄的增长,因为老龄化的发展,而不是反之亦然随着年龄下降的选择通常被视为老化的发展的原因。但是选择也可以随年龄的增加而增加的。然后会发生什么呢?不过这项工作表明,老龄化的发展,所以选择随着年龄的下降。
- 斯特凡诺Giaimo
- &阿恩特劳尔森
-
文章
|开放获取 的进化机制,在微生物产生表型异质性
的进化机制,在微生物产生表型异质性在微生物,细胞内人口经常显示极端表型变异与不同的机制用于确定如何分配不同的表型。本研究使用理论模型研究的相对优势两种主要机制,协调和随机的决心,在分割的劳动力数量和助手之间的微生物。
- 人亚历山大•库珀
- ,明刘
- &斯图亚特·安德鲁西
-
文章
|开放获取 指导微生物群落的生态学进化动力学改善人工选择
指导微生物群落的生态学进化动力学改善人工选择有效的微生物群落人工选择已被证明具有挑战性。在这个仿真研究中,作者探讨如何提高选择效果通过增加社区功能遗传实验可实现扰动策略。
- 李谢
- &署名为寿
-
文章
|开放获取 单细胞基因组学揭示plastid-lacking Picozoa是红藻的近亲
单细胞基因组学揭示plastid-lacking Picozoa是红藻的近亲的起源主要叶绿体Archaeplastida给了真核生物的祖先光合能力。本研究利用单细胞基因组学和phylogenomics推断的进化起源plastid-lacking门Picozoa,一群海洋微生物异养真核生物,这表明它们属于Archaeplastida和改变我们对质体演化的理解。
- 马克斯·e·肖恩
- ,瓦西里•诉Zlatogursky
- &法比安Burki
-
文章
|开放获取 秀丽隐杆线虫喂蛋黄年轻的一种原始的哺乳
秀丽隐杆线虫喂蛋黄年轻的一种原始的哺乳目前还不清楚为什么秀丽隐杆线虫继续生产大量繁殖后蛋黄。在这里作者表明,后生育期秀丽隐杆线虫母亲发泄蛋黄支持子女的成长,作为一种原始的哺乳。
- 船底座c·克恩
- ,StJohn汤森
- &大卫的宝石
-
评论文章
|开放获取 种子银行的原则和复杂性从休眠的出现
种子银行的原则和复杂性从休眠的出现种子银行时生成个人进入休眠状态,形成了一种现象,在不同的类群,但那是在干细胞还发现,大脑和肿瘤。在这里,列侬等人合成种子银行的基本理论和复杂的模式的出现和动力学数学和生命科学。
- 杰伊·t·列侬
- ,弗兰克窝荷兰人
- &Jochen Blath
-
文章
|开放获取 表观遗传记忆和传染病的进化
表观遗传记忆和传染病的进化一些传染病的毒性似乎依赖于宿主感染来自性,以及当前主机。在这里,麦克劳德等人开发一个流行病学模型探讨毒性当病原体的进化可以保留以前的主机的表观遗传的记忆。
- 大卫·v·麦克劳德
- ,Geoff野生
- &旧金山Ubeda
-
文章
|开放获取 快速和强大的自然选择的放大器
快速和强大的自然选择的放大器人口结构可以影响的概率和时间固定新的突变体。这里,Tkadlec等人证明数学结构,增加固定概率一定缓慢的固定,而且确定放大结构以最小的减少固定时间。
- 约瑟夫Tkadlec
- ,Andreas Pavlogiannis
- &马丁·a·诺瓦克
-
文章
|开放获取 无知之幕可以促进社会公平的哺乳动物
无知之幕可以促进社会公平的哺乳动物模糊知识的个人收益理论上个人可以维持公平合作小组。实验表明,野外,合作繁殖不确定的条纹猫鼬的亲属关系分配产后护理,减少不平等在后代中,显示一个典型的道德哲学的概念可以应用在生物系统。
- h·h·马歇尔
- ,r·a·约翰斯通
- &m·a .斜面
-
文章
|开放获取 灵长类动物的长寿和不变的老化速度的假设
灵长类动物的长寿和不变的老化速度的假设不变的老化速度的假设表明,老龄化的速度往往是不断在物种。这里,Colchero等人找到支持假说在灵长类动物,包括人类,表明生物老化的速度限制。
- 费尔南多Colchero
- ,若泽•曼努埃尔•Aburto
- &苏珊·c·爱尔
-
的角度来看
|开放获取 对一个工程进化论
对一个工程进化论有效的生物工程需要进化的确认及其在设计过程中考虑。在这个角度看,作者evotype原因有关的概念和形成自然和改造的生物系统的进化潜力。
- 西缅d城堡
- ,克莱尔·s·格里尔生家族的
- &托马斯·e·Gorochowski
-
文章
|开放获取 愿望动态生成健壮的异质人群的预测
愿望动态生成健壮的异质人群的预测社会互动的结果依赖于个体拥有的信息类型,以及它是如何用于决策。在这里,周等人发现自我评价为基础的决策规则导致进化的结果,是健壮的不同人口结构和自我评价的方法。
- Lei周
- ,吴本
- &王长
-
文章
|开放获取 共同进化过渡从对立到共生解释为选择对手的假设
共同进化过渡从对立到共生解释为选择对手的假设虽然有强有力的证据表明,许多从对立共生进化,如何或为何仍不清楚。结合理论研究和基于数据模型揭示了如何共生进化没有非常紧密的主人忠诚以及生态环境如何影响进化的结果,反之亦然。
- 克里斯托弗·a·约翰逊
- ,戈登·p·史密斯
- &瑞吉斯Ferriere
-
文章
|开放获取 氧抑制宏观的多细胞生物
氧抑制宏观的多细胞生物多细胞生物的进化被假设是由氧含量上升。通过实验进化和建模,Bozdag等人证明地球的第一个氧化会强烈限制,不推广,多细胞生物的进化。
- g . Ozan Bozdag
- ,埃里克·利比
- &威廉·c··拉特克利夫称
-
文章
|开放获取 分子进化和净化选择随着年龄的下降
分子进化和净化选择随着年龄的下降进化理论的一个基本原则是自然选择的力量较弱的特征表示在晚年相对于早期特征表达。在这里,作者发现强烈的分子进化和一致的模式反映出这一原则在四种动物,包括人类。
- 常德程
- &马克·柯克帕特里克
-
文章
|开放获取 进化的运动骨架安乐蜥蜴反映生态机会和惯性系统之间的相互作用
进化的运动骨架安乐蜥蜴反映生态机会和惯性系统之间的相互作用已经提出生态机会和表型模块化促进适应性辐射。菲娜等人证明安乐蜥蜴进化而来的一个新的模块化结构岛适应辐射,但是这种模块化多样化时,并没有产生相同的极端安乐回到大陆。
- 娜塔莉菲娜
- ,Illiam s c·杰克逊
- &托拜厄斯ul
-
文章
|开放获取 尽管在动态网络是会传染的
尽管在动态网络是会传染的是恶意的行为伤害演员和目标,它是具有挑战性的理解如何适应这些行为。这里富尔克等人表明,尽管可以通过反馈青睐与网络结构,同时创建相关和反关联行为的相互作用。
- 扎卡里·富尔克
- ,帕特里克forb
- &Christoph Riedl
-
评论
|开放获取 增加我们的能力来预测当代进化
增加我们的能力来预测当代进化经典的辩论有关科学家可以预测进化程度获得了新的紧迫感环境变化迫使物种适应或面临灭绝的风险。我们强调我们的预测能力进化是如何受到数据的限制,导致贫困的理解确定的自然选择。然后我们强调这些数据限制可以减少与可行的实证工作涉及的组合方法。
- 帕特里克•Nosil
- ,塞缪尔·m·Flaxman
- &撒迦利亚Gompert
-
文章
|开放获取 针锋相对的进化在细菌通过VI型分泌系统
针锋相对的进化在细菌通过VI型分泌系统博弈论社会进化的理解作出了巨大贡献。在一个优雅的结合实验测试和造型,这项研究表明,当细菌面对激烈的竞争,重复比一个针锋相对的回应袭击的报复。
- 威廉•p•j•史密斯
- ,Maj Brodmann
- &凯文·r·福斯特
-
文章
|开放获取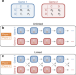 发展多渠道合作游戏
发展多渠道合作游戏大多数进化博弈理论侧重于孤立的游戏。这里,唐纳修等人提出了一个通用框架,“多通道游戏”中,个人参与的一组平行游戏伙伴,并展示了这种并行交互支持互惠的进化在游戏。
- 凯特·多纳休
- ,奥利弗·p·豪泽
- &基督教Hilbe
-
文章
|开放获取 进化的多面手抗除草剂混合显示阻力的一种权衡管理
进化的多面手抗除草剂混合显示阻力的一种权衡管理混合物的抗生素或杀虫剂可以帮助减少阻力单个化合物的演化。这里,Comont blackgrass等人表明,一个重要的农业杂草,减少除草剂混合做专门的阻力,而是可以选择广义抗性机制。
- 大卫Comont
- ,克劳迪娅·劳
- &保罗冰原
-
文章
|开放获取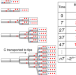 的机械解释过渡到简单的多细胞真菌
的机械解释过渡到简单的多细胞真菌多细胞生物在进化过程中是一个重大的转变。在这里,作者用一个模型表明,单细胞细菌相比,多细胞真菌可以更迅速殖民固定,营养价值资源因为菌丝体的胞外酶提供更大或更持久的好处生物。
- 卢克·l·m·希顿
- ,尼克·s·琼斯
- &马克·d·弗里克
-
文章
|开放获取 孩子在女性的准军事组织志愿者在第二次世界大战加速繁殖计划
孩子在女性的准军事组织志愿者在第二次世界大战加速繁殖计划生活史理论预测,女性将调整生育时机以应对环境挑战。这里作者表明,年轻女孩暴露于更高的死亡率在战争期间分娩,比同龄人更经常不暴露于这些条件。
- 罗伯特•林奇
- ,Virpi Lummaa
- &约翰Loehr
-
文章
|开放获取 湍流相干结构和早期生活在柯尔莫哥洛夫规模
湍流相干结构和早期生活在柯尔莫哥洛夫规模生命起源的模型通常需要一个机制来新兴人口结构。这里,克里格等人发展空间产生湍流流动模型表明,相干结构在水生环境中提供了隔间,促进生命的起源。
- 麦迪逊美国Krieger
- ,山姆西奈
- &马丁·a·诺瓦克
-
文章
|开放获取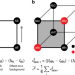 最低上位插值sequence-function关系
最低上位插值sequence-function关系高通量组合突变分析是有用的,屏幕的功能不同的序列,但他们并不详尽。这里,周和McCandlish开发一种方法推导出这种失踪genotype-phenotype数据基于推断最上位sequence-function关系。
- Juannan周
- &David m . McCandlish
-
文章
|开放获取 测量单在人体组织细胞分裂多区测序数据
测量单在人体组织细胞分裂多区测序数据量化体细胞在癌症和健康组织进化过程是一个挑战。在这里,作者使用单一时间点多区取样的癌症和正常组织,结合进化理论,量化体内突变每细胞分裂和细胞存活率。
- 本杰明·沃纳
- ,杰克的情况
- &安德里亚Sottoriva
-
文章
|开放获取 进化与环境反馈的游戏
进化与环境反馈的游戏战略游戏的回报往往取决于环境的状态,进而可以影响游戏策略。这里,Tilman等人开发一个通用的框架建模战略游戏环境反馈和分析案例研究从决策、生态学和经济学。
- 安德鲁·r·Tilman
- ,约书亚·b·普罗金
- &Erol Akcay
-
文章
|开放获取 得失推动原核生物基因同源重组障碍和加速基因组序列散度
得失推动原核生物基因同源重组障碍和加速基因组序列散度很大一部分细菌和古菌的分子进化发生通过基因得失。这里Iranzo等人开发一个数学模型来解释观察到的差异基因序列进化的模式与内容进化同源重组的结果。
- Jaime Iranzo
- ,尤里。狼
- &伊塔玛塞拉
-
文章
|开放获取 适应性维护议会的基因
适应性维护议会的基因“议会的基因”假说表明,自私基因元素将被抑制,保持中和相等的其他基因的传播。在这里,作者发现支持这一假设用数学模型来探索一系列不同的场景。
- 托马斯·w·斯科特
- &斯图亚特·a .西
-
文章
|开放获取 共同进化的男性和女性择偶可以破坏生殖隔离
共同进化的男性和女性择偶可以破坏生殖隔离配偶选择的模型主要集中在女性择偶的影响生殖隔离。在这里,Aubier等人开发一个人口之间的共同进化遗传模型女性和男性择偶,这会引起人口选型和随机交配之间摆动。
- 托马斯·g·Aubier
- ,汉娜合欢树
- &马修Joron
-
文章
|开放获取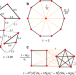 在等温图进化游戏
在等温图进化游戏人口的空间结构是经常合作的进化的关键。这里,艾伦和他的同事们表明,当空间结构由一个等温图,表示有效数字的邻居每个决定是否合作发展。
- 本杰明·艾伦
- ,伽柏Lippner
- &马丁·a·诺瓦克
-
文章
|开放获取 逐步转变背后的进化趋势哺乳动物脊柱形态的复杂性
逐步转变背后的进化趋势哺乳动物脊柱形态的复杂性哺乳动物在进化过程中脊柱变得更加复杂。这里,琼斯和他的同事们使用系统建模表明,这种复杂性增加逐步转变可能由适应有氧能力的提高。
- 卡特里娜·e·琼斯
- ,肯尼斯•d•Angielczyk
- &斯蒂芬妮·e·皮尔斯
 交互共存生态型之间在长期进化出现在权衡
交互共存生态型之间在长期进化出现在权衡