
文章
|开放获取
-
-
文章|
 E3连接酶适配器小脑靶向c端环亚胺去克隆
E3连接酶适配器小脑靶向c端环亚胺去克隆c端环酰亚胺是一种生理脱子,能够使泛素E3连接酶适配蛋白小脑蛋白靶向底物进行降解。
- 日本米酒川
- ,霍普·a·弗拉克斯曼
- &Christina M. Woo
-
文章
|开放获取 PRDM16稳定对米色脂肪生物形成的翻译后控制
PRDM16稳定对米色脂肪生物形成的翻译后控制泛素E3连接酶CUL2-APPBP2通过催化PRDM16在米色脂肪中的多泛素化来确定PRDM16蛋白的稳定性。
- 羌王
- ,Huixia李
- &shigo Kajimura
-
文章
|开放获取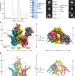 过氧化物酶体泛素连接酶复合物形成逆转录易位通道
过氧化物酶体泛素连接酶复合物形成逆转录易位通道膜嵌入的泛素连接酶复合物的冷冻电子显微镜结构揭示了其作为一个反向易位通道的功能,用于将移动受体从过氧化物酶体中穿梭出来。
- Peiqiang冯
- ,Xudong吴
- &汤姆·a·拉波波特
-
文章
|开放获取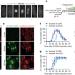 真核细胞周期的核心控制原理
真核细胞周期的核心控制原理核心细胞周期在很大程度上是由增加的总CDK活性以及CDKs启动DNA复制和有丝分裂的底物特异性的微小差异所驱动的。
- Souradeep巴苏
- ,杰西卡·格林伍德
- &保罗的护士
-
-
文章|
 ubr1介导N-degron多泛素化的结构研究
ubr1介导N-degron多泛素化的结构研究Ubr1与Ubc2、泛素和两个N-degron多肽的复合物结构揭示了Ubc2结合区域和Ubr1上的受体泛素结合环,为Ubr1催化泛素化的起始和延伸步骤提供了机制上的洞察。
- 人锅
- ,青云郑
- &Minglei赵
-
文章|
 志贺氏杆菌通过精氨酸adp核糖化caspase-11来避免焦亡
志贺氏杆菌通过精氨酸adp核糖化caspase-11来避免焦亡本研究报告了一种新的翻译后修饰,称为ADP核糖化,它是由酶介导的志贺氏杆菌并使caspase-4和caspase-11的胞质LPS感应通路失活。
- 张梓琳李
- ,王刘
- &冯邵
-
文章|
 BRCA1-BARD1核小体识别和泛素化机制
BRCA1-BARD1核小体识别和泛素化机制作者阐明了泛素连接酶复合物BRCA1-BARD1的泛素化特异性和募集到染色质内受损DNA以促进同源重组的机制。
- 气胡
- ,玛丽亚·维多利亚·博图扬
- &乔治Mer
-
文章|
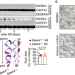 MARK4通过微管脱氨作用控制缺血性心力衰竭
MARK4通过微管脱氨作用控制缺血性心力衰竭MARK4通过促进MAP4磷酸化来调节心肌细胞的收缩能力,从而促进VASH2进入微管进行α-微管蛋白的脱酪化;急性心肌梗死后MARK4缺乏限制左室射血分数的降低。
- 西安昱
- ,小陈
- &李宣
-
文章|
 细菌感染时RNF213对脂多糖的泛素化作用
细菌感染时RNF213对脂多糖的泛素化作用在沙门氏菌E3泛素连接酶RNF213对哺乳动物细胞质的入侵、非蛋白质底物(细菌脂多糖的脂质a部分)的泛素化标志着细菌作为抗菌自噬的载体。
- 埃尔斯杰·g·奥滕
- ,艾玛·沃纳
- &Felix Randow
-
文章|
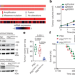 NSD3组蛋白甲基化活性升高导致鳞状细胞肺癌
NSD3组蛋白甲基化活性升高导致鳞状细胞肺癌组蛋白H3K36甲基转移酶NSD3与常见的8p11-12染色体扩增相关,是肺鳞状细胞癌的致癌驱动因素。
- 帮派元
- ,娜塔莎·m·弗洛雷斯
- &或Gozani
-
文章|
 H1组蛋白通过局部染色质压实控制表观遗传景观
H1组蛋白通过局部染色质压实控制表观遗传景观使用条件三敲除小鼠品系的实验表明,组蛋白H1通过控制染色质压实、基因组结构和组蛋白甲基化来调节染色质域的活性。
- 迈克尔·a·威尔考森
- ,肖恩·e·希尔顿
- &Arthur I. Skoultchi
-
文章|
 剪接因子YBX1介导持久性JAK2突变的肿瘤
剪接因子YBX1介导持久性JAK2突变的肿瘤抑制YBX1, Janus激酶JAK2的下游靶点,使骨髓增生性肿瘤细胞对JAK敏感,并可能为根除人类造血癌症中的此类细胞提供一种手段。
- Ashok Kumar Jayavelu
- ,蒂娜·M. Schnöder
- &弗洛里安·h·海德尔
-
-
文章|
 二聚体质量控制的结构基础
二聚体质量控制的结构基础对E3型泛素连接酶SCF-FBXL17二聚体质量控制的结构研究表明,SCF-FBXL17对异常复合物形成的选择性是基于对相互作用域的形状和互补性的识别。
- 以利亚·l·梅纳
- ,据Jevtić
- &迈克尔强奸
-
文章|
 包膜蛋白泛素化驱动寨卡病毒进入和发病机制
包膜蛋白泛素化驱动寨卡病毒进入和发病机制E3泛素连接酶TRIM7多泛素化寨卡病毒的包膜蛋白,添加lys63连接的多泛素链,与宿主细胞的TIM1受体相互作用,增强病毒的进入和复制。
- 玛丽亚·吉拉尔多
- ,Hongjie夏
- &里卡多Rajsbaum
-
文章|
 多(UG)尾rna在基因组保护和表观遗传中的作用
多(UG)尾rna在基因组保护和表观遗传中的作用在秀丽隐杆线虫,核糖核苷酸转移酶RDE-3将交替的尿苷和鸟苷核糖核苷酸添加到RNA的3 '端,这是RNA干扰的关键步骤,从而实现表观遗传秀丽隐杆线虫生殖系。
- Aditi舒克拉
- ,珍妮严
- &斯科特•肯尼迪
-
文章|
 配体诱导的BIK1单泛素化调控植物免疫
配体诱导的BIK1单泛素化调控植物免疫通过植物细胞检测微生物相关配体激活信号级联,其中激酶BIK1被单泛素化,从FLS2-BAK1复合物中释放,并通过内吞作用内化。
- 西域马
- ,卢卡斯·a·n·克劳斯
- &荔波山
-
文章|
 晚期促进复合物控制的基因表达和细胞特性
晚期促进复合物控制的基因表达和细胞特性WDR5和TBP将后期促进复合物招募到有丝分裂中的特定转录起始位点,启动一种依赖泛素的机制,通过连接基因表达和细胞分裂来保持细胞特性。
- 尤金哦
- ,凯文·g·马克
- &迈克尔强奸
-
文章|
 HPF1完成PARP活性位点,用于DNA损伤诱导的adp核糖化
HPF1完成PARP活性位点,用于DNA损伤诱导的adp核糖化人类细胞DNA损伤后,HPF1与PARP1或PARP2结合形成的催化中心的组装对于蛋白质adp核基化至关重要。
- Marcin J. Suskiewicz
- ,Florian祖贝儿
- &伊凡Ahel
-
-
文章|
 宿主介导的分枝杆菌蛋白泛素化抑制免疫
宿主介导的分枝杆菌蛋白泛素化抑制免疫结核分枝杆菌通过分枝杆菌蛋白的宿主介导泛素化抑制宿主细胞产生炎症细胞因子,增强宿主信号抑制剂与另一信号分子的相互作用。
- 林王
- ,Juehui吴
- &Baoxue通用电气
-
文章|
 伴侣蛋白对哺乳动物细胞α-突触核蛋白的调控
伴侣蛋白对哺乳动物细胞α-突触核蛋白的调控伴侣蛋白与α-突触核蛋白中的典型基序相互作用,这可以通过α-突触核蛋白Tyr39位点的磷酸化来阻止,而抑制这种相互作用会导致α-突触核蛋白定位于线粒体并形成聚集体。
- Björn M. Burmann
- ,胡安·a·杰雷兹
- &塞巴斯蒂安•希勒
-
文章|
 动态谱系启动是由ERK直接增强子调控驱动的
动态谱系启动是由ERK直接增强子调控驱动的ERK通过辅助因子和RNA聚合酶从多能性到早期分化增强子的选择性再分配可逆地调节胚胎干细胞转录,同时使转录因子与其增强子结合,从而保持可塑性。
- 威廉·b·汉密尔顿
- ,Yaron Mosesson
- &约书亚·m·布里克曼
-
文章|
 范可尼贫血单泛素连接酶复合物的结构
范可尼贫血单泛素连接酶复合物的结构利用冷冻电子显微镜和质谱确定的多蛋白Fanconi贫血核心复合体的结构表明,该复合体采用了扩展的不对称结构,突出了环指结构域的结构和功能不对称。
- Shabih沙基尔
- ,Eeson Rajendra
- &洛莉·a·帕斯莫尔
-
-
信|
 使用Ub-clipping对泛素链架构的洞察
使用Ub-clipping对泛素链架构的洞察泛素分子内的酶裂解,随后进行定量质谱分析,简化了复杂的泛素链,并实现了多泛素结构的映射。
- Kirby N. Swatek
- ,乔安妮·亚瑟
- &大卫Komander
-
信|
 P38γ在细胞周期进程和肝脏肿瘤发生中至关重要
P38γ在细胞周期进程和肝脏肿瘤发生中至关重要应激激活激酶p38γ在调节进入细胞周期中起作用;在肝脏再生过程中可诱导细胞增殖,促进肝细胞癌的发展。
- 安东尼娅Tomas-Loba
- ,Elisa Manieri
- &瓜达卢佩Sabio
-
信|
 组蛋白5 -羟色胺化是一种允许修饰,可增强TFIID与H3K4me3的结合
组蛋白5 -羟色胺化是一种允许修饰,可增强TFIID与H3K4me3的结合在富含血清素的组织中,组织转谷氨酰胺酶2能够将5 -羟色胺连接到组蛋白H3中的谷氨酰胺残基上;这种修饰介导这些组织中的允许基因表达。
- 洛娜·a·法雷利
- ,罗伯特·e·汤普森
- &伊恩迷宫
-
信|
 pkg1修饰的TSC2调节mTORC1活性以对抗心脏不良应激
pkg1修饰的TSC2调节mTORC1活性以对抗心脏不良应激TSC2中两个相邻丝氨酸残基之一的磷酸化对于pkg1介导的小鼠心脏抗压力过载保护是必需的,也是充分的;这些丝氨酸残基为胁迫刺激下mTORC1活性的双向调控提供了遗传工具。
- 马克·j·拉尼克
- ,克里斯汀·m·科克南-西蒙
- &大卫·a·卡斯
-
信|
 SETD3是一种肌动蛋白组氨酸甲基转移酶,可预防原发性难产
SETD3是一种肌动蛋白组氨酸甲基转移酶,可预防原发性难产SETD3使哺乳动物肌动蛋白His73位点甲基化,SETD3缺乏损害刺激诱导的原代人子宫平滑肌细胞收缩,导致小鼠母体难产。
- 亚历克斯·w·威尔金森
- ,乔纳森Diep
- &或Gozani
-
信|
 代谢重编程年代-亚硝基辅酶a还原酶系统保护肾脏免受损伤
代谢重编程年代-亚硝基辅酶a还原酶系统保护肾脏免受损伤AKR1A1-regulated蛋白质S -亚硝基化通过pkm2介导的代谢重编程保护肾脏免受损伤。
- 花林周
- ,Rongli张
- &乔纳森·s·斯坦勒
-
信|
 钯介导的酶活化提示糖发生的多阶段启动
钯介导的酶活化提示糖发生的多阶段启动由糖原素启动的糖发生机制包括三个不同的动力学阶段,最后一个阶段涉及精炼过程,其中只有葡萄糖作为底物被耐受。
- 马修·k·比尔雅德
- ,亨利·j·贝利
- &本杰明·g·戴维斯
-
文章|
 无膜细胞器有丝分裂中激酶控制的相变
无膜细胞器有丝分裂中激酶控制的相变双特异性激酶DYRK3作为一个中心的“溶解酶”,在有丝分裂过程中介导几种类型的无膜细胞器的相变。
- Arpan Kumar Rai
- ,Jia-Xuan陈
- &卢卡斯Pelkmans
-
信|
 OTULIN通过去泛素化LUBAC限制细胞死亡和炎症
OTULIN通过去泛素化LUBAC限制细胞死亡和炎症OTULIN去除LUBAC沉积的泛素链,通过阻止其自身泛素化来促进LUBAC活性,从而支持正常小鼠胚胎发育并防止成年小鼠促炎细胞死亡。
- 克劳斯Heger (
- ,凯瑟琳·e·威克利夫
- &Vishva M. Dixit
-
信|
 PINK1激活帕金蛋白的机制
PINK1激活帕金蛋白的机制全长人帕金蛋白的结构质谱和被激活的帕金蛋白核的结构揭示了PINK1激活帕金蛋白过程中涉及的大规模结构域重排。
- 克里斯蒂娜Gladkova
- ,莎拉·l·马斯林
- &大卫Komander
-
-
-
-
信|
 LUBAC通过防止细胞死亡和使造血成为胚胎发生所必需的
LUBAC通过防止细胞死亡和使造血成为胚胎发生所必需的LUBAC泛素连接酶复合物的HOIL-1组分是LUBAC活性所必需的,LUBAC活性通过防止异常tnfr1介导的内皮细胞死亡和ripk1介导的造血缺陷来防止胚胎发生过程中的致命性。
- Nieves peltz
- ,莫里斯·达尔德人的
- &亨宁查克
-
信|
 基于活性的E3连接酶分析揭示了具有酯化活性的E3连接酶
基于活性的E3连接酶分析揭示了具有酯化活性的E3连接酶E3泛素连接酶MYCBP2的非赖氨酸泛素化活性通过活性分析确定;对MYCBP2的生化和结构分析为其机制和特异性奠定了基础。
- Kuan-Chuan Pao
- ,尼古拉·t·伍德
- &Satpal Virdee
-
信|
 蛋白质组氨酸磷酸酶LHPP是一种肿瘤抑制剂
蛋白质组氨酸磷酸酶LHPP是一种肿瘤抑制剂组氨酸磷酸酶LHPP是一种新型的肿瘤抑制因子,其表达的降低导致整体组氨酸磷酸化和肝细胞癌的增加。
- Sravanth K. Hindupur
- ,马可Colombi
- &迈克尔·n·霍尔
-
信|
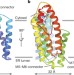 真核生物膜内RAS甲基转移酶ICMT的原子结构
真核生物膜内RAS甲基转移酶ICMT的原子结构整体膜蛋白异丙基半胱氨酸羧甲基转移酶的x射线结构表明了它识别水溶性和膜结合的反应物以催化RAS和其他CAAX蛋白在膜-细胞质界面的甲基化的机制。
- 梅琳达·m·戴弗
- ,琳恩没效果
- &斯蒂芬·b·朗
-
信|
 KAT2A结合α-KGDH复合物起组蛋白H3琥珀酰转移酶的作用
KAT2A结合α-KGDH复合物起组蛋白H3琥珀酰转移酶的作用组蛋白乙酰转移酶KAT2A(也称为GCN5)也可以催化组蛋白琥珀酰化,α-KGDH复合物提供了琥珀酰辅酶a的局部来源。
- Yugang王
- ,郭玉松
- &Zhimin陆
-
信|
 启动子结合的METTL3通过m维持髓系白血病6依赖a的平移控制
启动子结合的METTL3通过m维持髓系白血病6依赖a的平移控制甲基转移酶METTL3促进急性髓系白血病(AML)的白血病状态通过催化m6通过在aml相关基因的转录起始位点募集的RNA修饰。
- Isaia巴比里
- ,Konstantinos Tzelepis
- &托尼Kouzarides
-
信|
 Cyclin D-CDK4激酶通过cullin 3-SPOP稳定PD-L1控制癌症免疫监测
Cyclin D-CDK4激酶通过cullin 3-SPOP稳定PD-L1控制癌症免疫监测PD-L1是抗癌免疫治疗靶点PD-1的配体,通过细胞周期蛋白D-CDK4 /cullin 3-SPOP轴的多泛素化负调控,PD-1阻断治疗与CDK4/6抑制剂联合使用可提高小鼠的生存率。
- Jinfang张
- ,夏Bu
- &Wenyi魏
-
信|
 泛素依赖信号轴特异性alkbh介导的DNA脱烷基修复
泛素依赖信号轴特异性alkbh介导的DNA脱烷基修复人类细胞中检测由烷基化引起的DNA损伤的信号机制包括泛素依赖性的烷基化修复复合物ASCC募集到损伤附近,并与转录和剪接因子共定位。
- 约书亚·r·布里克纳
- ,詹妮弗·m·索尔
- &尼玛Mosammaparast
-
文章|
 PINK1与底物泛素的复合物结构
PINK1与底物泛素的复合物结构利用纳米体稳定瞬时蛋白激酶-底物复合物提供了关于帕金森病相关蛋白激酶PINK1如何磷酸化泛素的分子细节,并提出了新的药理学策略。
- 亚历山大·f·舒伯特
- ,克里斯蒂娜Gladkova
- &大卫Komander
-
文章|
 选择性小分子抑制剂抑制USP7的分子基础
选择性小分子抑制剂抑制USP7的分子基础经鉴定,小分子可以抑制泛素特异性蛋白酶USP7,具有高亲和力和共晶体结构所解释的特异性,并被证明可以减少小鼠的肿瘤生长。
- 安德鲁·特恩布尔
- ,名字埃尼迪斯
- &大卫Komander
 聚合活化罐基酶的结构基础
聚合活化罐基酶的结构基础 小G蛋白香叶基化监测细胞内脂质
小G蛋白香叶基化监测细胞内脂质 STAT3棕榈酰化循环促进TH辨证与结肠炎
STAT3棕榈酰化循环促进TH辨证与结肠炎 相分离组织自噬体形成的部位
相分离组织自噬体形成的部位 组蛋白乳糖化对基因表达的代谢调控
组蛋白乳糖化对基因表达的代谢调控 磷酸核糖-泛素化机制的单一介导军团菌效应
磷酸核糖-泛素化机制的单一介导军团菌效应 磷酸核糖连接丝氨酸泛素化的催化和功能研究
磷酸核糖连接丝氨酸泛素化的催化和功能研究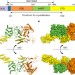 泛素修饰的结构基础军团菌效应SdeA
泛素修饰的结构基础军团菌效应SdeA