
文章
|开放获取
-
-
文章|
 Bestrophin-2和谷氨酰胺合成酶形成谷氨酸释放复合物
Bestrophin-2和谷氨酰胺合成酶形成谷氨酸释放复合物电生理、结构和生化研究表明,bestrophin-2阴离子通道对谷氨酸的渗透性不对称,并与谷氨酰胺合成酶形成协同机制释放谷氨酸。
- 亚伦·p·奥吉
- ,块玉
- &婷婷杨
-
文章
|开放获取 肌动蛋白丝组装及老化的结构基础
肌动蛋白丝组装及老化的结构基础骨骼f -肌动蛋白的冷冻电子显微镜结构显示,溶剂驱动的重排控制着肌动蛋白丝的组装和老化,在药物和小分子成像和治疗的设计中具有潜在的应用价值。
- Wout Oosterheert
- ,Björn U.克林
- &Stefan Raunser
-
文章
|开放获取 弯曲力和核苷酸状态共同调节f -肌动蛋白结构
弯曲力和核苷酸状态共同调节f -肌动蛋白结构肌动蛋白的核苷酸状态调节弯曲力引起的f -肌动蛋白结构转变。
- 马修·j·雷诺兹
- ,卡拉Hachicho
- &Gregory M. Alushin
-
-
文章|
 多通道膜蛋白的膜内伴侣机制
多通道膜蛋白的膜内伴侣机制在多通道膜蛋白生物发生过程中,中间产物的生化和结构分析显示了膜内伴侣如何引导新生膜蛋白进入一个半封闭的充满脂质的腔,在那里它们被正确插入和折叠。
- 卢卡Smalinskaitė
- ,金敏京
- &Ramanujan S. Hegde
-
-
文章
|开放获取 TMC-1复合体的结构阐明了机械感觉转导
TMC-1复合体的结构阐明了机械感觉转导天然跨膜通道样蛋白1 (TMC-1)机械感觉转导通道复合物的结构研究秀丽隐杆线虫揭示亚基组成和蛋白质-膜相互作用在机械力转换为离子通道活性的作用。
- Hanbin宋
- ,莎拉克拉克
- &Eric Gouaux
-
文章
|开放获取 ω - rna和靶DNA配合物中OMEGA - nickase IsrB的结构
ω - rna和靶DNA配合物中OMEGA - nickase IsrB的结构低温电子显微结构d . thermocuniculiIsrB蛋白及其同源的不饱和rna和目标DNA的复合物表明rna主导的IsrB效应复合物与蛋白质主导的Cas9效应复合物共享一个共同的支架。
- Seiichi Hirano
- ,Kalli卡柏
- &冯张
-
文章|
 与核小体结合的NuA4乙酰转移酶复合物的结构
与核小体结合的NuA4乙酰转移酶复合物的结构来自NuA4的冷冻电镜结构酿酒酵母与核小体结合说明了NuA4是如何组装的,并为组蛋白乙酰转移酶对核小体识别和转录共激活的机制提供了见解。
- 瞿温驯
- ,Kangjing陈
- &诸城陈
-
文章
|开放获取 细菌细胞内原子细节的翻译动态可视化
细菌细胞内原子细节的翻译动态可视化冷冻电子断层扫描技术用于揭示翻译核糖体的结构动力学和功能多样性肺炎支原体,提供了深入了解细胞内的翻译延伸周期以及抗生素如何重塑它。
- 梁雪
- ,Swantje楞次
- &茱莉亚Mahamid
-
-
文章
|开放获取 小麦抗性小体定义了免疫受体通道的共同原理
小麦抗性小体定义了免疫受体通道的共同原理植物受体结构的进化保护允许小麦和大麦核苷酸结合富亮氨酸重复受体(NLRs)的新变体的产生,这些变体识别小麦茎锈病病原的AvrSr35,支持基于结构的NLRs工程用于作物改良的原理的证明。
- 亚历山大Forderer
- ,Ertong李
- &Jijie柴
-
文章|
 微生物糖苷水解酶激活植物受体样蛋白
微生物糖苷水解酶激活植物受体样蛋白一项聚焦于植物免疫的结构分析揭示了含有lrr的受体样蛋白如何识别致病配体并因此被激活,数据表明这些蛋白质通过两种不同的机制靶向病原体。
- 悦的太阳
- ,王阎
- &Jijie柴
-
-
文章|
 呼吸复合体I的普遍偶联机制
呼吸复合体I的普遍偶联机制的冷冻电子显微镜研究大肠杆菌我提出了一种耦合质子转移和静电相互作用的保守机制,导致质子只在远端NuoL亚基从复合物中喷射出来。
- Vladyslav克拉夫丘克
- ,奥尔加佩特洛娃
- &列昂尼德•Sazanov
-
-
文章|
 微管上dynein-dynactin的结构显示串联适配器结合
微管上dynein-dynactin的结构显示串联适配器结合完整的dynein-dynactin复合物的结构及其与微管和负载适配器的相互作用使用冷冻电子显微镜可见。
- 萨米Chaaban
- &安德鲁·卡特
-
文章|
 人对甲氨蝶呤的识别降低了叶酸载体SLC19A1
人对甲氨蝶呤的识别降低了叶酸载体SLC19A1Cryo-EM结构可以深入了解抗叶酸甲氨蝶呤(一种化疗药物)是如何被减少的叶酸载体识别的。
- 尼古拉斯·j·赖特
- ,贾斯汀·g·费多尔
- &Seok-Yong李
-
文章|
 在光捕获和光保护状态下藻胆体的结构
在光捕获和光保护状态下藻胆体的结构低温电子显微结构集胞藻属藻胆体-单独和结合橙色类胡萝卜素蛋白-揭示了关于控制蓝藻光收集的生物物理基础的详细信息。
- María Agustina Domínguez-Martín
- ,保罗·v·绍尔
- &谢丽尔·a·科菲尔德
-
文章
|开放获取 Cas9的r环形成及构象活化机制
Cas9的r环形成及构象活化机制冷冻电子显微镜结构酿脓链球菌Cas9在多种DNA结合状态下的发现为研究Cas9被目标DNA激活的机制提供了新的思路。
- 马丁Pacesa
- ,Luuk Loeff
- &马丁Jinek
-
文章
|开放获取 AAA+ atp酶介导的RuvAB-Holliday结分支迁移机制
AAA+ atp酶介导的RuvAB-Holliday结分支迁移机制以多种构象捕获的atp水解RuvAB复合物的结构提供了在DNA重组过程中协调atp酶和运动活性的机制见解。
- 雅罗西克瓦尔德
- ,德克Fahrenkamp
- &托马斯·c·马洛维斯
-
文章|
 选择性TnsC招募增强rna引导转位的保真度
选择性TnsC招募增强rna引导转位的保真度crispr相关转座子利用校对检查点,通过级联复合物特异性招募TnsC低聚物,确保高保真地选择DNA插入的基因组位点。
- 弗洛里安·t·霍夫曼
- ,Minjoo金
- &塞缪尔·h·斯滕伯格
-
文章|
 环核苷酸诱导的螺旋结构激活TIR免疫效应
环核苷酸诱导的螺旋结构激活TIR免疫效应细菌抗病毒防御系统产生一种环三腺苷酸,与TIR - saved效应物结合,诱导形成一种超螺旋结构,相邻的TIR结构域组织成一个活性位点,从而导致NAD+退化。
- 流行Hogrel
- ,阿比公会
- &马尔科姆·f·怀特
-
文章
|开放获取 SARS-CoV-2的RNA封盖机制
SARS-CoV-2的RNA封盖机制SARS-CoV-2 RNA 5 '帽的重构揭示了SARS-CoV-2 RNA基因组帽的非常规机制,为开发治疗COVID-19的抗病毒药物提供了新的靶点。
- 吉娜·j·帕克
- ,亚当Osinski
- &文森特·s·塔利亚布拉奇
-
文章|
 激素和抗体介导的促甲状腺素受体的激活
激素和抗体介导的促甲状腺素受体的激活促甲状腺激素和自身抗体M22使促甲状腺素受体的胞外结构域呈直立的活性构象,揭示了糖蛋白激素受体的普遍激活机制,为Graves病、甲状腺功能减退症和桥本病的发生提供了分子基础。
- 贾段
- ,Peiyu徐
- &许耀威
-
文章|
 促甲状腺素受体激素作用的自身抗体模拟
促甲状腺素受体激素作用的自身抗体模拟甲状腺激素受体的冷冻电子显微镜结构揭示了Graves病患者自身抗体激活受体的基础。
- 布莱恩·浮士德
- ,Christian B. Billesbølle
- &Aashish Manglik
-
文章
|开放获取 巨型噬菌体核壳的结构与自组装
巨型噬菌体核壳的结构与自组装细菌被巨型噬菌体201phi2-1感染时形成的类核腔室由噬菌体蛋白chimallin组成,在体外可以自组装成封闭的腔室。
- 托马斯·g·劳克林
- ,阿玛深
- &伊丽莎白的别墅
-
文章
|开放获取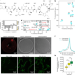 Teixobactin通过双管齐下地攻击细胞包膜来杀死细菌
Teixobactin通过双管齐下地攻击细胞包膜来杀死细菌结合上述方法,揭示了抗生素teixobactin的作用机理。
- 节奏舒克拉
- ,弗朗西斯卡Lavore
- &马库斯Weingarth
-
文章
|开放获取 生长素识别和外排的结构见解拟南芥PIN1
生长素识别和外排的结构见解拟南芥PIN1的结构拟南芥生长素输出体PIN1处于载波状态,与天然生长素结合或与抑制剂结合,为PIN家族转运蛋白介导的生长素极性转运机制提供了见解。
- Zhisen杨
- ,静夏
- &Linfeng太阳
-
文章|
 的结构和机制拟南芥生长素转运蛋白PIN3
的结构和机制拟南芥生长素转运蛋白PIN3拟南芥PIN3结构揭示了吲哚-3-乙酸转运和抑制生长素极性转运的分子机制N1-naphthylphthalamic酸。
- 小南南苏
- ,朱爱琴告诉我们
- &Jiangtao郭
-
文章|
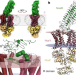 一种氧化亚氮还原酶组装机械的分子相互作用
一种氧化亚氮还原酶组装机械的分子相互作用参与氧化亚氮生产和功能的细菌蛋白质机制的冷冻电子显微镜结构提供了这种酶的组装途径和铜运输机制的见解。
- Christoph穆勒
- ,林张
- &奥利弗Einsle
-
文章
|开放获取 四醚类脂合酶的发现、结构及作用机理
四醚类脂合酶的发现、结构及作用机理在Methanocaldococcus jannaschii,激进分子年代-腺苷甲硫氨酸酶催化双植物链的形成。
- 科迪·t·劳埃德
- ,大卫·f·艾维格
- &Squire J. Booker
-
文章
|开放获取 IL-17配体-受体轴的组织结构原则
IL-17配体-受体轴的组织结构原则IL-25-IL-17RB-IL-17RA和IL-17A-IL-17RC-IL-17RA复合物的冷冻电镜结构显示尖端到尖端的结构,这是IL-17受体家族的一个关键组织原理。
- 史蒂文·c·威尔逊
- ,Nathanael A. Caveney
- &K.克里斯托弗·加西亚
-
文章|
 膜锚定HDCR纳米线驱动氢动力CO2固定
膜锚定HDCR纳米线驱动氢动力CO2固定丝状氢依赖CO的冷冻电镜结构2还原酶(HDCR)Thermoanaerobacter kivui结合酶分析和原位冷冻电子断层扫描技术,研究了HDCR的高催化活性。
- 海尔格·m·迪特里希
- ,里卡多·d·Righetto
- &简·m·舒勒
-
文章
|开放获取 活性细菌TIR-STING纤维复合物的低温电镜结构
活性细菌TIR-STING纤维复合物的低温电镜结构通过对细菌STING活化的结构分析,确定了STING长丝形成和TIR效应域在抗噬菌体信号传导中的激活的分子基础。
- 本杰明·r·莫尔豪斯
- ,叶志强
- &菲利普·j·克兰祖施
-
文章
|开放获取 古风陪护引子毛里自隐成超弹性之字形弹簧
古风陪护引子毛里自隐成超弹性之字形弹簧多药耐药医院病原菌Csu菌毛鲍曼不动杆菌组装成一个超薄的锯齿形结构,由一个固定机制,提供高机械稳定性和超弹性的毛。
- 纳塔莉亚Pakharukova
- ,亨利Malmi
- &安东·v·扎维亚洛夫
-
文章|
 的结构四膜虫端粒酶结合的CST与聚合酶α-引物酶
的结构四膜虫端粒酶结合的CST与聚合酶α-引物酶冷冻电子显微镜结构四膜虫thermophila端粒酶结合的Ctc1-Stn1-Ten1和DNA聚合酶α -引物酶为端粒复制和维持的分子机制提供了新的思路。
- 么他
- ,他的歌
- &朱莉Feigon
-
文章
|开放获取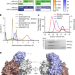 MRAS-SHOC2-PP1C磷酸酶复合物的结构
MRAS-SHOC2-PP1C磷酸酶复合物的结构MRAS-SHOC2-PP1C复合物的结构支持ras驱动的RAF激活多分子模型,其中单个RAS-GTP分子招募RAF - 14-3-3和SHOC2-PP1C来激活下游通路。
- 扎卡里·j·豪斯曼
- ,米歇尔Fodor
- &丹尼尔·a·金
-
-
-
文章|
 端粒模板结合的人cst - pol α引物酶复合物的结构
端粒模板结合的人cst - pol α引物酶复合物的结构结构分析表明单链DNA结合辅助蛋白复合物CST如何物理组织人类DNA聚合酶-α -引物酶复合体,以在端粒复制期间有效地合成引物。
- Qixiang他
- ,Xiuhua林
- &林慈济
-
-
文章
|开放获取 SHOC2调控RAS信号的结构基础
SHOC2调控RAS信号的结构基础SHOC2-PP1C - mras复合物的冷冻电子显微镜结构、分子动力学和生化分析证明了复合物的形成对RAS - gtp的依赖性,并确定了RAS亚型对SHOC2-PP1C偏好的决定因素以及复合物对RAF去磷酸化的特异性。
- Nicholas p.d. Liau
- ,马修·c·约翰逊
- &dina Sudhamsu
-
文章
|开放获取 对dsRNA处理的结构见解果蝇Dicer-2-Loqs-PD
对dsRNA处理的结构见解果蝇Dicer-2-Loqs-PDDcr-2 - Loqs-PD复合物在处理双链RNA (dsRNA)底物时的结构阐明了Dcr-2和Loqs-PD之间的相互作用,并表明Dcr-2在dsRNA处理周期中经历了大量的构象变化。
- Shichen苏
- ,贾汪
- &金标马
-
文章
|开放获取 植物PIN-FORMED生长素转运体的结构与机制
植物PIN-FORMED生长素转运体的结构与机制结构和生物物理分析拟南芥生长素转运蛋白PIN8揭示了PIN转运蛋白使用升降机机制输出生长素。
- 坚林雄
- ,米凯尔温克勒
- &Bjørn Panyella Pedersen
-
文章
|开放获取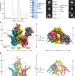 过氧化物酶体泛素连接酶复合物形成逆转录易位通道
过氧化物酶体泛素连接酶复合物形成逆转录易位通道膜嵌入的泛素连接酶复合物的冷冻电子显微镜结构揭示了其作为一个反向易位通道的功能,用于将移动受体从过氧化物酶体中穿梭出来。
- Peiqiang冯
- ,Xudong吴
- &汤姆·a·拉波波特
-
文章
|开放获取 Dicer-2-R2D2异源二聚体与小RNA双工结合的结构
Dicer-2-R2D2异源二聚体与小RNA双工结合的结构冷冻电子显微镜结构果蝇含有和不含有小干扰RNA的Dicer-2-R2D2复合物揭示了RNA如何以正确的方向呈现给Argonaute,以进行病毒基因沉默。
- Sonomi山口
- ,Masahiro Naganuma
- &Osamu Nureki
-
文章|
 eIF5B和eIF1A重新定位启动器tRNA,允许核糖体亚基连接
eIF5B和eIF1A重新定位启动器tRNA,允许核糖体亚基连接使用单分子光谱和结构研究来检查eIF1A和eIF5B与人类翻译起始复合物的关联动力学,以及它们在将tRNA呈现给复合物以启动翻译中的作用。
- Christopher P. Lapointe
- ,罗斯林格罗斯
- &约瑟夫·d·普格利西
-
文章
|开放获取 IV型分泌系统的冷冻电镜结构
IV型分泌系统的冷冻电镜结构由质粒R388编码并包含92个多肽的2.8兆达尔顿细菌IV型分泌系统的冷冻电子显微镜结构为毛毛组装的阶梯式机制提供了深入的了解。
- 凯文·梅斯
- ,阿比纳夫·k·瓦达克帕特
- &加布里埃尔Waksman
 SEA复合物的冷冻电镜结构
SEA复合物的冷冻电镜结构 埃博拉病毒聚合酶复合物的结构
埃博拉病毒聚合酶复合物的结构 定向甲壳素生物合成的结构基础
定向甲壳素生物合成的结构基础 人端粒染色质的柱状结构
人端粒染色质的柱状结构 SHOC2-MRAS-PP1C全磷酸酶复合物的结构功能分析
SHOC2-MRAS-PP1C全磷酸酶复合物的结构功能分析 豪猪介导的Wnt酰化机制及抑制
豪猪介导的Wnt酰化机制及抑制 营养传感中心GATOR2的结构
营养传感中心GATOR2的结构